Transcriber's Note: Original spelling and punctuation have been
retained. In particular, both Eutainia and Eutaenia are used in
the original, as are both pickeringi and pickeringii.
University of Kansas Publications
Museum of Natural History
Volume 13, No. 5, pp. 289-308, 4 figs.
February 10, 1961
Occurrence of the Garter Snake,
Thamnophis sirtalis,
in the Great Plains and Rocky Mountains
BY
HENRY S. FITCH AND T. PAUL MASLIN
University of Kansas
Lawrence
1961
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Robert W. Wilson
Volume 13, No. 5, pp. 289-308, 4 figs.
Published February 10, 1961
University of Kansas
Lawrence, Kansas
PRINTED IN
THE STATE PRINTING PLANT
TOPEKA, KANSAS
1961
Occurrence of the Garter Snake,
Thamnophis sirtalis,
in the Great Plains and Rocky Mountains
BY
HENRY S. FITCH AND T. PAUL MASLIN
Introduction
The common garter snake (Thamnophis sirtalis) has by far the
most extensive geographic range of any North American reptile,
covering most of the continental United States from the Atlantic to
the Pacific and from south of the Mexican boundary far north into
Canada and southeastern Alaska. Of the several recognized subspecies,
the eastern T. s. sirtalis has the most extensive range, but
that of T. s. parietalis in the region between the Mississippi River
and the Rocky Mountains is almost as large. The more western
T. s. fitchi occurring from the Oregon and California coasts east
through the northern Great Basin, has the third largest range, while
the far western subspecies pickeringi, concinnus,
infernalis and tetrataenia,
and the Texan T. s. annectens all have relatively small ranges.
Since the publication of Ruthven's revision of the genus Thamnophis
more than 50 years ago, little attention has been devoted to
the study of this widespread and variable species, except in the
Pacific Coast states (Van Denburgh, 1918; Fitch, 1941; Fox, 1951).
However, Brown (1950) described the new subspecies annectens
in eastern Texas, and many local studies have helped to clarify the
distribution of the species in the eastern part of the continent and
to define the zone of intergradation between the subspecies sirtalis
and parietalis. In our study attention has been focused upon
parietalis
in an attempt to determine its western limits and its relationships
to the subspecies that replace it farther west.
Taxonomic History
Thamnophis sirtalis parietalis Say was described (as Coluber
parietalis) in 1823 from a specimen obtained in what is now Washington
County, Nebraska, on the west side of the Missouri River
three miles upstream from the mouth of Boyer's River [Iowa], or
approximately eight miles north of Omaha. Although the type locality
was unequivocally stated in the original description, Nebraska
was not mentioned since the state was not yet in existence. Because
[pg 292]
the mouth of Boyer's River, the landmark by means of which the
type locality is defined, is in Iowa, the impression has been imparted
that the type locality itself is in Iowa (Schmidt, 1953:175), and to
our knowledge the type locality has never been associated with Nebraska
in the literature.
Like all the more western subspecies, parietalis is strikingly different
from typical sirtalis in having conspicuous red markings. The
relationship between the two was early recognized. Several of the
other subspecies were originally described as distinct species. Coluber
infernalis Blainville, 1835; Tropidonotus concinnus Hallowell,
1852; Eutainia pickeringi Baird and Girard, 1853; and others now
considered synonyms eventually came to be recognized as conspecific
with Thamnophis sirtalis. Ruthven (1908:166-173) allocated
all western sirtalis to either parietalis or concinnus, the
latter
including the populations of the northwest coast in Oregon, Washington
and British Columbia.
Subsequent more detailed studies by later workers with more
abundant material led to the recognition of some subspecies that
Ruthven thought invalid and led to the resurrection of some names
that he had placed in synonomy. Van Denburgh and Slevin (1918:198)
recognized infernalis as the subspecies occurring over most
of California and southern Oregon, differing from more northern
populations in having more numerous ventrals and caudals and a
paler ground color. Fitch (1941:575) revived the name pickeringii
for a melanistic population of western Washington and southwestern
British Columbia, restricting the name concinnus to a red-headed
and melanistic population of northwestern Oregon, and restricting
the name infernalis to a pale-colored population in the coastal strip
of California.
These changes left most of the populations formerly included in
concinnus and infernalis without a name, and Fitch (op.
cit.) revived
Thamnophis sirtalis tetrataenia (Cope) to apply to them. However,
Fox (1951:257) demonstrated that the type of T. s. tetrataenia came
from the San Francisco peninsula (rather than from "Pit River, California"
as erroneously stated in the original description) and that
the name was applicable to a localized peninsular population rather
than to the wide-ranging far western subspecies, which he named
T. s. fitchi. The range of fitchi includes California west of the
Colorado
and Mohave deserts (except for the narrow strip of coast
occupied by infernalis and tetrataenia), Oregon except the northwestern
[pg 293]
part, Washington east of the Cascade Range, most of British
Columbia, extreme southeastern Alaska (occurring farther north
than any other terrestrial reptile of North America) and parts of
Idaho.
Neither Fox (1951) nor Fitch (1941) defined the eastern limits
of fitchi or discussed its relationship to the subspecies
parietalis.
Wright and Wright (1957:849) stated: "Fitch ... did not
even mention the big scrap basket form parietalis, from which he
pulled T. s. fitchi (old tetrataenia). That comparison remains to
be
made, and the east boundary of fitchi and the west boundary of
parietalis are still nebulous." We have undertaken to define better
than has been done before the ranges of parietalis and fitchi and
to list the diagnostic characters separating these two subspecies.
Freshly collected material of both has been compared. At the time
of his 1941 revision the senior author had never seen a live or recently
preserved specimen of parietalis.
Discontinuity of Range
Wherever it occurs at all, the common garter snake is usually
abundant. Because of its diurnal habits and the concentration of
its populations along watercourses, it is not likely to be overlooked.
There are few, if any, remaining large areas in the United States
where herpetologists have not carried on field work. It may be
anticipated that certain rare and secretive species will still be found
far from any known stations of occurrence, and seeming gaps in the
ranges of these species will eventually be filled. But for the common
garter snake the negative evidence provided by the lack of records
from extensive areas should be taken into account in mapping the
range.
Most large collections of garter snakes contain misidentified specimens.
The diagnostic differences in color and pattern are often
obscured, especially if the specimens are poorly preserved. Many
specimens deviate from the scalation typical of the form they represent,
and key out to other species. Isolated records should therefore
be accepted with caution. A case in point is Colorado University
Museum No. 46, from Buford, Rio Blanco County, Colorado, originally
identified by Cockerell (1910:131) as Thamnophis sirtalis
parietalis. This specimen, and another, now lost, from Meeker in the
same county seemingly served as the basis for mapping the range
of sirtalis across the western half of Colorado, for there seem to be
[pg 294]
no other records from this part of the state. However, a re-examination
of the specimen from Buford shows it to be an atypical individual
of another species, T. elegans vagrans. A specimen of T.
radix haydeni (Col. U. Mus. No. 3165) was the basis for Maslin's
(1959:53) record of parietalis in Baca County on the north fork of
the Cimarron River in southeastern Colorado. Brown (1950:203)
has mentioned the difficulty of defining the range of sirtalis in the
southern Great Plains because of misidentifications of the similar
T. radix.
The range of the common garter snake has never been adequately
mapped in the Rocky Mountain and Great Basin states. Recent general
works (Smith, 1956:291; Wright and Wright 1957:834; Stebbins
1954:505; Conant 1958:328) which have shown maps of the over-all
range of sirtalis, differ sharply as to the extent of its distribution in
Texas, New Mexico and Arizona, but all show its distribution as
continuous over the more northern Great Basin and Rocky Mountain
states. However, specimens and specific locality records from this
extensive area seem to be scarce and some are based on early collections
of doubtful provenance. Throughout this region the low
rainfall, fluctuating and uncertain water supply, and general lack of
mesic vegetation along many of the streams render the habitat rather
hostile to garter snakes in general. Thamnophis elegans vagrans,
highly adapted to conditions in this region and generally distributed
over it, doubtless offers intensive competition to the species sirtalis
wherever they overlap and perhaps constitutes a limiting factor for
sirtalis in some drainage basins.
Convincing records of sirtalis are lacking from all of Colorado—except
for those in the drainage basins of the South Platte, and the
Río Grande east of the Continental Divide—from the eastern half
of Utah (east of the Wasatch Range), from New Mexico except for
the Río Grande drainage (with one record each for the Canadian
and Pecos river drainages), from southwestern Wyoming (at least
that part in the Colorado River drainage basin), from the western
half of Oklahoma, and from Texas, except the eastern and extreme
western and northern parts. The species occurs in Nevada only
near that state's western and northern boundaries. The range is
therefore much different than it has been depicted heretofore, with
the populations living east of the Continental Divide widely separated
from those to the west for the entire length of the Rocky
Mountains south of the Yellowstone National Park region. The populations
of northern Utah, southern Idaho, and Nevada, which have
[pg 295]
been considered parietalis are thus far removed from the main
population of that subspecies to the east and are isolated from them
by the barrier of the Continental Divide and arid regions farther
west.
Although some of the records published for Thamnophis sirtalis
are erroneous, being based on misidentifications of other species,
various outlying records, including those in western Kansas, the
Panhandle of Texas, and southeastern New Mexico probably represent
localized relict populations that have survived from a time when
the species was more generally distributed in this region. The population
of T. sirtalis in the Río Grande drainage of New Mexico is
geographically isolated and remote from other populations of the
species. Except for a few isolated and highly localized populations
the species is absent from the Republican, Smoky Hill, Arkansas,
Cimarron, Canadian, Red, Brazos, Colorado and Pecos rivers and
their tributaries west of the one hundredth meridian in the arid High
Plains.
Streams in this region of High Plains are in most instances unsuitable
habitats because they are in eroded channels, have a variable
and uncertain water supply, and have poorly developed riparian
communities. The marsh and wet meadow habitat preferred by
sirtalis in most parts of its range is almost absent. T. radix and
T.
marcianus, well adapted to conditions in this region, perhaps provide
competition that is limiting to T. sirtalis. However, several
well-isolated
populations of sirtalis have survived as relicts in the southern
Great Plains, presumably from a time several thousand years ago
when mesic conditions were more prevalent, perhaps in an early
postglacial stage.
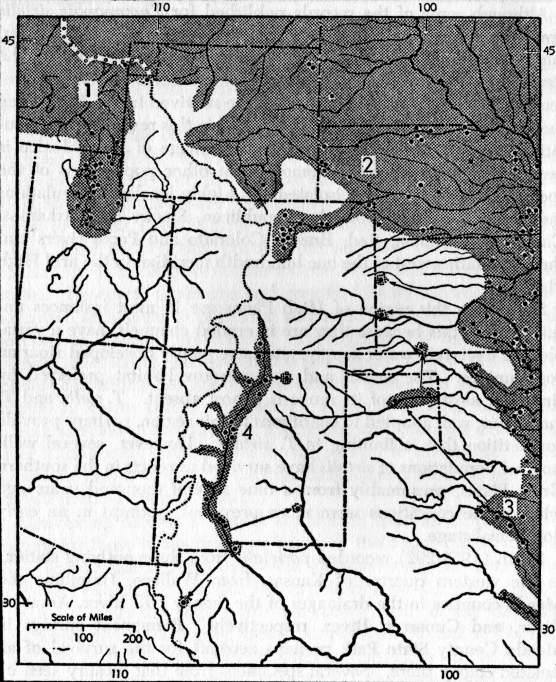
Fig. 1. Map of a part of the United States in the
region of the Great Plains and
Rocky Mountains, and adjacent northwestern Mexico showing supposed range
(shaded) and localities of authenticated occurrence (dots) of Thamnophis
sirtalis. 1. T. s. fitchi, 2. T. s. parietalis, 3. T. s.
annectens, 4. T. s. ornata. Records
from Idaho and Wyoming are based on specimens in the University of
Kansas Museum of Natural History collection. Other records are based on
Woodbury (1931) for Utah, Hudson (1942) for Nebraska, Maslin (1959) for
Colorado, Smith (1956) for Kansas, R. G. Webb (MS) for Oklahoma, Brown
(1950) and Fouquette and Lindsay (1955) for Texas, Cope (1900), Van Denburgh
(1924), Little and Keller (1937) for New Mexico, and Smith and Taylor
(1945) for Mexico.
Smith (1956:292) recorded parietalis from three outlying stations
in the western quarter of Kansas, from Wallace, Hamilton and
Meade counties in the drainages of the Smoky Hill River, Arkansas
River, and Cimarron River, respectively. Permanent springs in
Meade County State Park perhaps account for the survival of an
isolated colony there. Several specimens from that locality seen by
Fitch in August, 1960, when recently collected by a University of
Michigan field party, seemed to be of the Texas subspecies annectens,
as their dorsal stripes were reddish orange, and markings on
the dorsolateral area were pale yellow rather than red. Specimens
from the Texas Panhandle, from Hemphill County (Brown, 1950:207)
and nine miles east of Stinnet, Hutchison County (Fouquette
and Lindsay, 1955:417) likewise are most nearly like annectens
[pg 296]
judging from the authors' descriptions. The specimens from nine
miles east of Stinnet averaged large; the two largest would have
attained or slightly exceeded four feet in length if they had had complete
[pg 297]
tails. No sirtalis so long as four feet has been recorded
elsewhere.
Records are lacking from the drainages of the Republican, North
Canadian, Brazos and Colorado River drainages in the High Plains,
but possibly isolated populations occur in some of these also. The
only record from the Pecos River drainage is that of Bundy (1951:314)
from Wade's Swamp near Artesia, Eddy County, New Mexico.
This locality is separated by some 140 miles from any other known
station of occurrence.
From extreme southern Colorado south across New Mexico to the
Mexican border T. sirtalis occurs in continuous or nearly continuous
populations in the Río Grande Valley, and has been recorded from
many localities. It has been recorded from relatively few localities
of tributary streams (Los Pinos, Abiqui, Santa Fe) all near the main
valley. There is one record from the Ocate River, a headwaters
tributary of the Canadian River, in the Sangre de Cristo Mountains
near other localities in the Río Grande drainage. The southwestern-most
known locality of occurrence is Casas Grandes in the Mexican
state of Chihuahua some 130 miles southwest of El Paso, Texas, and
near the Continental Divide. The Río Casas Grandes must have
once been a tributary of the Río Grande, but now its desert drainage
basin is isolated.
Re-description of a Subspecies from New Mexico
Most specimens of a population of sirtalis occurring in New Mexico
are recognizably different from most specimens of other populations.
This New Mexican population is therefore here recognized
as a distinct subspecies:
Thamnophis sirtalis ornata Baird
Eutaenia ornata Baird, 1859:16.
Eutaenia sirtalis dorsalis Cope, 1900:1076.
Thamnophis sirtalis parietalis (part) Van Denburgh, 1924:222.
Type.—U. S. Nat. Mus. No. 960, obtained at El Paso, Texas, at some time in
the eighteen fifties by Col. J. D. Graham.
Range.—Río Grande and vicinity, from Conejos and Costilla counties in extreme
south-central Colorado south across New Mexico to Mexican border.
Records from neighboring drainage systems, Casas Grandes in Chihuahua and
Artesia and Ocate River in New Mexico, probably also pertain to ornata.
Description.—A specimen in the University of New Mexico Natural History
Museum (E. D. Flaherty No. 560, obtained one mile west and one-half mile
south of Isleta, Bernalillo County, New Mexico, on May 31, 1959) was described
as follows while its colors were still but little altered by preservatives:
Top of head olive, supralabials pale gray, edged with black posteriorly; chin
milky white, with dark edges posteriorly on fifth, sixth and seventh infralabials;
[pg 298]
dorsal stripe yellow; including middorsal row of scales and little more than adjacent
half of row on either side of it; dorsolateral area olive-brown with row of
black spots on its lower half, these spots elliptical, averaging about size of one
scale on anterior part of body, smaller posteriorly; adjacent spots separated by
interspaces of approximately their own length, irregular black markings on upper
half of dorsolateral area not forming definite spots but fused longitudinally to
form continuous black border to dorsal stripe; crescent-shaped red markings in
areas between scale rows three to nine, these markings invading edges of scales,
and themselves having ill-defined edges blending into the darker ground color;
lateral stripe pale, yellowish gray, limited to scale rows two and three for most
of its length, but including rows four and five in neck region; row of irregular
black marks low on each side, with each mark centering on anterior part of
lower half of scale of first row but overlapping onto corners of adjacent ventrals;
approximately every other scale of first row so marked; ventral surface pale,
suffused with bluish tint; most of ventrals marked on anterior edges with pair of
semicircular black spots, each situated about two-thirds of distance from mid-line
to lateral edge of ventral; these marks diminishing in size and finally disappearing
on posterior part of body; ventral surface otherwise immaculate.
Lepidosis normal for genus and species, with preoculars single on each side,
supralabials 7-7, infralabials 8-8, ventrals 159, anal entire, subcaudals 77 (including
terminal spine), paired except for second, third and fourth; scale rows
19 from neck slightly beyond mid-body, fifth on left side ending opposite 86th
ventral; length from snout to vent 670 mm., tail 202 mm.
Comparisons.—From T. s. parietalis, T. s. ornata differs in its consistently pale
ground color, olive instead of dark brown or black. In respect to color-pattern
ornata stands in approximately the same relation to parietalis as, farther west,
T. s. infernalis, a pale subspecies of the California coast, stands in relation to
T. s. fitchi. Nevertheless, fitchi consistently has a dark ground color, whereas
parietalis is highly variable, and the color of an occasional specimen (for example
KU 17032 from Douglas County, Kansas) matches ornata in olive coloration.
These unusually pale specimens of parietalis differ from ornata in not
having a continuous black edge along each side of dorsal stripe; black pigment
of this area is concentrated into rows of spots alternating with those of lower
series. From T. s. infernalis, ornata differs in having paired black spots on the
ventrals and in having more than three series of red crescents on dorsolateral
area of each side.
Remarks.—The type of ornata seems to have been lost, and the
available information concerning it is far from satisfactory. In the
original description, Baird listed three specimens, purportedly from
"Indianola, Texas" (J. H. Clark, 438), from the Río Grande, Texas
(J. H. Clark, 768), and from near San Antonio, Texas (Dr. Kennerley,
no number). None of these three specimens could have been
ornata as conceived of by us because all were collected outside the
geographic range of ornata. However, there was also included a
plate with a drawing of a specimen and a reference to an earlier
paper (Baird and Girard, 1853) in which a specimen obtained by
Col. Graham "Between San Antonio and El Paso" was described.
Smith and Brown (1946:72) have presented evidence that this specimen
[pg 299]
figured (rather than any of the three specifically mentioned)
served as a basis for the plate, and they therefore considered it to
be the holotype of ornata, even though Baird referred this specimen
to "Eutaenia parietalis Say" in the same paper (1859) in which the
original description of ornata was published. Cope (1900:1079)
listed under Eutaenia sirtalis parietalis a specimen, U. S. Nat. Mus.
No. 960, from El Paso, obtained by Col. Graham, and referred to it
as a type (without specifying of what it was the type). Smith and
Brown (loc. cit.) interpreted this statement by Cope as further evidence
that the specimen in question should be considered the type
of ornata, and they restricted the type locality, originally stated as
"between San Antonio and El Paso" to "El Paso." Actually all valid
records of the species sirtalis from the vicinity of the Río Grande are
from the El Paso region or from farther north.
It is with some misgivings that we herewith accept the interpretation
proposed by Smith and Brown regarding the applicability of
the name ornata and the designation by these authors of the now
missing specimen from the region of El Paso as the holotype of that
form. The evidence linking the name ornata with the New Mexican
subspecies is tenuous; there is some doubt as to the provenance of
U. S. Nat. Mus. No. 960 (the supposed type), and even more doubt
as to whether this is the specimen depicted in the plate that formed
part of the original description.
Cope (1900:1076) recognized as a distinct subspecies, Eutaenia
sirtalis dorsalis, the same population that we recognize herein as
T. s. ornata, and Smith (1942:98) considered the name dorsalis to
be a synonym of T. s. parietalis. However, it is almost certain that
both authors misapplied the name, since the type of Baird's and
Girard's (1853:31) Eutainia dorsalis was obtained
in Coahuila, Mexico,
between Monclova and the Río Grande, far south of the known
range limits of T. sirtalis in Texas. The description does not fit T.
sirtalis and almost certainly pertains to another species.
Specimens examined.—Univ. of Kansas Mus. Nat. Hist. (hereafter abbreviated
to "KU") Nos. 5479 to 5497, from the north end of Elephant Butte
Reservoir, Sierra County, New Mexico, and 8592 and 8593 from near Las Lunas,
Valencia County, New Mexico; Univ. of New Mexico Mus. Nos. 571 and 572
(J. S. Findley) from 2 miles west and 1/4 mile north of Albuquerque, Bernalillo
County, New Mexico, and No. 4021 (E. D. Flaherty) from 1 mile west and 1/2
mile south of Isleta, Bernalillo County, New Mexico.
Description of T. s. parietalis
From most of the vast area occupied by parietalis, material has
not been available to us, and our concept of this subspecies is based
chiefly on specimens and living material from Kansas and northeastern
[pg 300]
Colorado. A total of 520 live parietalis has been examined
from the University of Kansas Natural History Reservation some 130
miles south and a little east of the type locality in Nebraska. These
probably differ but little from typical specimens. The range of individual
variation in pattern is especially notable. In those from
the Reservation, the ground color varies from dull olive-brown to
almost jet black. The markings on the dorsolateral area vary in
color, in shade and in extent. These marks are chiefly confined to
the skin between the scales of rows three to nine. Although most
typically these marks are of some shade of red (hence the name
"red-sided garter snake"), they may be pale buff, or pale greenish
yellow, or may even have a bluish tint. In approximately ten per
cent of the specimens from the Reservation there is no red at all in
the pattern, which hence is similar to that of T. s. sirtalis in the
eastern
United States. Only a minority have all the dorsolateral marks
red, and in some of these specimens the marks higher on the sides
are progressively paler red, having a bleached out appearance. Most
typically the marks between rows three to six are some shade of red
while the smaller marks between rows six to nine are pale—yellowish,
greenish, or buffy. In some the pale area of the lateral stripe is
in varying degrees suffused with red, which may extend onto the
edges of the ventrals and even to the underside of the tail.
T. s. parietalis may be diagnosed, on the basis of these snakes from
northeastern Kansas, as follows: Size medium large (length 23.5 to
34.5, or, exceptionally 43.5 inches in adult males; 32.5 to 46.0 inches
in adult females), dorsolateral color olive to black. Approximately
every other scale of the third row is bordered above and anteriorly
by a crescent-shaped area of scarlet colored skin. Similar crescent-shaped
areas border the scales of the fourth and fifth rows and often
two adjacent crescents meet at the ends of an intervening scale and
fuse forming an H-shaped mark. Placed alternately with these markings
are similar but smaller crescent-shaped markings on the skin
of the upper half of the dorsolateral area on each side bordering
every other scale of the sixth, seventh and eighth rows. The crescents
of this upper series also may fuse to form series of H-shaped
markings alternating with those of the lower series. The dorsal stripe
is yellow with a faint dusky suffusion; it involves all of the middorsal
scale row and approximately the adjacent half of the row on either
side. The lateral stripe is faint, yellowish gray, chiefly on the upper
half of the second scale row, lower half of third, and the intervening
skin, and is often invaded or suffused by the red marks of the dorsolateral
area. The first scale row, adjacent corners of the ventrals,
[pg 301]
and lower half of the second scale row are suffused with dark pigment
and appear dusky, but this area is often marked with black,
setting off the paler area of the lateral stripe. The ventrals are dull,
whitish, faintly suffused with yellowish, greenish or bluish, each
ventral having a black dot usually of semicircular shape on its anterior
margin near the anterolateral corner.
Comparison of T. s. parietalis and T. s. fitchi
Like most widely ranging subspecies, parietalis and fitchi vary
geographically and local populations often are noticeably different
from typical material. It is possible that future revisors will recognize
additional subspecies, but in the variant populations known
to us the degree of differentiation is slight as compared, for instance,
with that in the subspecies of Thamnophis elegans. Scalation is
remarkably
uniform in all the subspecies of sirtalis, but coastal and
northern populations tend to have fewer ventrals and subcaudals
than do their counterparts farther inland and farther south. In their
geographic variation the ventrals and subcaudals follow clines, and
do not in themselves warrant subspecific divisions. Variation occurs
chiefly in the color and pattern including the intensity of dark pigmentation
of the dorsolateral area, head, ventral surface and lower
edge of the lateral stripe; in extent, position and shade of red or pale
colored markings on the dorsolateral area; in presence and extent of
reddish suffusion on the head, in the region of the lateral stripe, and
on the ventral surface of the tail. Most of these same characters
[pg 302]
vary within the subspecies fitchi, but the range of variation is
relatively
minor. Fitch (op. cit.:582-584) described typical populations
and also described briefly several small series from British Columbia,
Idaho, Oregon, and California which were not entirely typical. Most
[pg 303]
frequent variation was in heavy reddish suffusion on the sides of the
head not found in typical fitchi. In each local population of this
subspecies the characters seem to be remarkably uniform and stable.

Fig. 2. Diagrammatic drawing of pattern in stretched
skin of T. s. fitchi; the
pale markings on the black dorsolateral area are scarlet (× 2-1/2).

Fig. 3. Diagrammatic drawing of stretched skin of T.
s. parietalis; the scarlet
markings extend farther dorsally than in T. s. fitchi and black spots are
prominent
on the ventrals laterally. Some individuals of parietalis have much paler
ground color, resembling ornata except in minor details (× 2-1/2).

Fig. 4. Diagrammatic drawing of stretched skin of T.
s. ornata. The ground
color is like that of parietalis but paler with a continuous black area
bordering
the dorsal stripe (× 2-1/2).
T. s. parietalis differs from fitchi in several trenchant
characters,
and there are additional slight or average differences between the
two. In approximate order of their importance the differences are
as follows: 1) The red (or pale yellow or green or buffy) marks on
skin between the scales on the upper half of the dorsolateral area
(that is between the sixth and seventh, seventh and eighth and
eighth and ninth scale rows) present in parietalis are missing in
fitchi or are represented by only an occasional small fleck. 2) The
dorsolateral area is black or nearly so in fitchi but averages paler in
parietalis, in which a wide range of shades may be found from black
to olive brown. 3) The red of the dorsolateral area frequently invades
the lateral stripe, which sometimes is mostly red, and may
even invade the ventrals in parietalis, but in fitchi the red
marks are
usually confined to the dorsolateral area, and do not invade the
lateral stripe. 4) The prominent paired black dots or semicircular
marks on the anterior edge of each ventral in parietalis are largely
lacking in fitchi, which rarely has any dark marks on the ventral
surface. 5) The dorsal stripe consistently involves the middorsal
scale row and the adjacent half of the next row on each side, and is
bright yellow in fitchi, but in parietalis it may be slightly
wider, may
be duller with more dusky suffusion, and its edges may be less
sharply defined.
Intermediate and Atypical Populations
Of many specimens examined from eastern Oregon, Idaho, Utah,
Wyoming and Colorado, few were typical of either parietalis or
fitchi. Many were intermediate in some respects or showed a composite
of characters of the two subspecies. No well-defined belt of
intergradation exists, but the transition extends over more than a
thousand miles, with local populations somewhat isolated and
slightly differentiated along divergent lines. In view of this situation
some plausibility could be claimed for any of several dividing lines
between the subspecies. However, by far the most logical division
is the Continental Divide; south of the Teton Range it constitutes
a broad barrier separating eastern and western populations. Across
Montana and Canada also it constitutes a more or less formidable
barrier, with high altitudes and cold climates that probably are limiting
to garter snakes. With few exceptions the snakes from east of
the Continental Divide are more nearly like parietalis in the sum of
[pg 304]
their characters whereas those from west of the Divide are more
nearly like fitchi.
In the Teton Range and in Yellowstone National Park these garter
snakes occur in headwater streams up to the Continental Divide.
KU 27956 from Two Ocean Lake 3-1/2 miles northeast of Moran, Teton
County, Wyoming, agrees in its characters with fitchi, having the
red lateral marks small and inconspicuous, discernible only on the
anterior half of the body. The dorsolateral area is dark, almost
black. The ventrals lack dark markings.
In Utah, populations of sirtalis occur in the drainages of the Bear,
Weber and Sevier rivers and other smaller streams of the western
half of the state. Obviously the species invaded Utah from the
north, probably at a time when Lake Bonneville, the predecessor
of the present Great Salt Lake, drained into the Snake River of
Idaho. Van Denburgh and Slevin (1918:190) separated from their
western "concinnus" and "infernalis" and allocated to
parietalis the
populations of Utah and southeastern Idaho, but presumably these
authors were not familiar with typical parietalis of the Mid-west.
Subsequent authors (Wright and Wright, 1957:834; Stebbins, 1954:505;
Conant, 1958:328) have followed this arrangement. A re-examination
of specimens from Utah, including living individuals
collected at Bear Lake in the summer of 1959, indicates that they
should be assigned to fitchi rather than to parietalis.
Likewise various specimens from the drainage basin of the Snake
River in Idaho are predominantly fitchi in the sum of their characters,
although they differ from that subspecies in its most typical
form and resemble parietalis in some respects. KU 23133 from two
miles east of Notus, Canyon County, Idaho, has the red crescents
on the lower part of the sides (between scale rows six and seven)
consistently developed on the anterior half of the body. KU 21873,
a large female from Bannock County, Idaho, is exceptional in having
small lateral black spots on the ventrals, resembling parietalis most
closely in this respect. Also, it has the red lateral crescents unusually
well developed; the first three series are conspicuous, those
of the fourth series are consistently developed, and those of the fifth
series show occasionally.
Forty-five specimens in the University of Colorado Museum from
northwestern Colorado were subjected to pattern analysis. In three
specimens the dorsolateral black area between the dorsal stripe and
the lateral stripe on each side has no markings, and in eight others
there is only an occasional fleck or crescent on the skin between the
[pg 305]
sixth and seventh scale rows. All others have the normal complement
of dorsolateral crescents or flecks between the scales of rows
three and four, four and five, and five and six. But, these specimens
vary in extent of development of the crescents in the upper half of
the dorsolateral area on each side—between scale rows six and seven,
seven and eight, and eight and nine. Only six snakes show traces of
the crescents of the uppermost series (between scale rows eight and
nine). Development of these crescents is variable but in all the
specimens the crescents are confined to the anterior half of the body.
The crescents between rows six and seven and between seven and
eight are present in 20 specimens and in ten of these the crescents
are conspicuous and regularly arranged, often meeting and consequently
form H-shaped markings. In most of the snakes the crescents
are best developed in the second fifth of the body and disappear
posteriorly. In five of the twenty, crescents between rows six and
seven are fairly regular, but those between rows seven and eight are
few and appear only sporadically. In eight specimens there are no
crescents between either rows seven and eight or eight and nine. In
eight others the crescents between rows six and seven are likewise
absent, and only the crescents between rows three to six are present.
These specimens from Colorado also differ from typical parietalis
in having the black spots on the anterolateral edges of the ventrals
less developed. In three of the 45 these spots are lacking entirely
and in four others they are few and small. In the majority of specimens
the spots are from 1/4 to 1/5 the length of the ventrals. In approximately
one-third of the specimens the spots are absent posterior to
mid-body. In five specimens obtained at Sheridan Lake, Pennington
County, South Dakota, in the Black Hills in August, 1960, dorsolateral
areas are dark with red crescents small and inconspicuous,
and with black spots either lacking from the ventrals or only faintly
developed. In two specimens from Sundance, Crook County, northeastern
Wyoming, the red crescents are small and inconspicuous
also. In one of these specimens, KU 28028, small black spots are
present in the corners of the ventrals, but in the other, KU 23654, the
spots are absent.
In having the dorsolateral area consistently black, with the three
uppermost series of red crescents reduced or absent, and in having
the ventral black spots reduced or absent, these specimens from
Colorado, Wyoming, and South Dakota differ from more eastern and
more typical parietalis, and tend toward fitchi, even more
strongly
than some Idaho specimens tend toward parietalis. Nevertheless,
[pg 306]
all things considered, the Continental Divide is the most logical
boundary between the two subspecies, even though occasional individuals
and even local populations deviate from the general trend
of characters from east to west.
Acknowledgments
Dr. Doris M. Cochran of the United States National Museum kindly furnished
information concerning the type specimen of Eutainia dorsalis formerly in the
National Museum collection but now lost. Dr. James S. Findley of the University
of New Mexico and Dr. Ralph J. Raitt of New Mexico State University
contributed habitat notes and records of specimens and loaned us critical specimens
of T. sirtalis from New Mexico. Drs. George F. Baxter of the University
of Wyoming, John M. Legler of the University of Utah, and Wilmer W. Tanner
of Brigham Young University kindly provided us with information concerning
the specimens in the collections of their respective institutions, and their personal
observations concerning the distribution of garter snakes in their states.
Alice V. Fitch, Chester W. Fitch and Donald S. Fitch assisted in the collection
of fresh specimens in Oregon and Utah and the unsuccessful search of many
a mosquito-infested meadow in southern Wyoming and northwestern Colorado
in July, 1959. Dr. R. G. Webb made available his MS on reptiles of Oklahoma.
This taxonomic study of garter snakes originated as a by-product of the
senior author's study of ecology and economic bearing of snakes in the central
Plains Region of the United States, for which support received from the National
Science Foundation is gratefully acknowledged.
Literature Cited
Baird, S. F.
1859. Reptiles of the boundary. United States and Mexican Bound. Surv.,
2, 1-35, 41 pls.
Baird, S. F., and Girard, C.
1853. Catalogue of North American reptiles in the Museum of the Smithsonian
Institution. Smithson. Miscl. Col., part 1, Serpents., pp. xvi
+ 172.
Brown, B. C.
1950. An annotated check list of the reptiles and amphibians of Texas.
Baylor Univ. Studies, pp. xii + 259.
Bundy, R. E.
1951. New locality records of reptiles in New Mexico. Copeia, 1951 (4):314.
Cockerell, T. D. A.
1910. Zoology of Colorado. Univ. of Colorado, Boulder, vii + 262 pp.
Conant, R.
1958. A field guide to the reptiles and amphibians of the United States and
Canada east of the 100th Meridian. Houghton Mifflin Co., Boston,
xviii + 366 pp.
Cope, E. D.
1900. The crocodilians, lizards and snakes of North America. Rept. U. S.
Nat. Mus. for 1898, pp. 153-1270.
Fitch, H. S.
1941. Geographic variation in garter snakes of the species Thamnophis
sirtalis in the Pacific Coast region of North America. Amer. Midland
Nat., 26:570-592.
Fox, W.
1951. The status of the gartersnake, Thamnophis sirtalis tetrataenia.
Copeia, 1951:257-267.
Fouquette, M. J., and Lindsay, H. L., Jr.
1955. An ecological survey of reptiles in parts of northwestern Texas.
Texas Jour. Sci., 7(4):402-421.
Hudson, G. E.
1942. The amphibians and reptiles of Nebraska. Nebraska Conserv. Bull.
No. 24, pp. 1-146.
Little, E. L., Jr., and Keller, J. G.
1937. Amphibians and reptiles from the Jornada Experimental Range,
New Mexico. Copeia, 1937 (4):402-421.
Maslin, T. P.
1959. An annotated check list of the amphibians and reptiles of Colorado.
Univ. Colorado Studies, Ser. Biol. No. 6, 98 pp.
Ruthven, A. G.
1908. Variations and genetic relationships of the garter-snakes. Bull. U. S.
Nat. Mus., 61:xii + 201 pp.
Schmidt, K. P.
1953. A check list of North American amphibians and reptiles. Univ.
Chicago Press, vii + 280 pp.
Smith, H. M.
1942. The synonymy of the garter snakes (Thamnophis), with notes on
Mexican and Central American species. Zoologica, 27(17):97-123.
1956. Handbook of amphibians and reptiles of Kansas (2nd ed.). Univ.
Kansas Mus. Nat. Hist. Misc. Publ. No. 9, 356 pp.
Smith, H. M., and Brown, B. B.
1946. The identity of certain specific names in Thamnophis. Herpetologica,
3:71-72.
Smith, H. M., and Taylor, E. H.
1945. An annotated check list and key to the snakes of Mexico. Bull. U. S.
Nat. Mus., 187, 239 pp.
Stebbins, R. C.
1954. Amphibians and reptiles of western North America. McGraw-Hill
Book Co., Inc., xxiv + 528 pp.
Van Denburgh, J.
1924. Notes on the herpetology of New Mexico, with a list of the species
known from the state. Proc. California Acad. Sci., 4th ser., 13(12):189-250.
Van Denburgh, J., and Slevin, J.
1918. The garter-snakes of Western North America. Proc. California
Acad. Sci., 4th ser., 8:181-270.
Woodbury, A. M.
1931. A descriptive catalog of the reptiles of Utah. Univ. Utah Biol.
Ser., 1(4):1-129.
Wright, A. H., and Wright, A. A.
1957. Handbook of snakes of the United States and Canada. Comstock
Publ. Associates, Cornell Univ. Press, vol. 2, pp. i + ix and 565-1106.
Transmitted November 8, 1960.