British Museum (Natural History)
Ground Floor.
Transcriber’s Note:
New original cover art included with this eBook is granted to the public domain.
British Museum (Natural History)
Ground Floor.
One of the large north galleries approached from the Bird Gallery is devoted to the exhibition of the extensive Class of Mollusca. Specimens of the types of all the principal divisions of this Class are exhibited, either entire and preserved in spirit, or as models. However, as not the animals, but their shells have always been a favourite object of study, and a popular source of pleasure to collectors, the exhibition of the species of shells has been made as complete as the space of this Gallery admitted. It has thus proved adequate for the requirements of the majority of visitors and students who consult this Collection. A separate series of British Shells is exhibited in some small table-cases along the west wall.
This Gallery also contains the exhibited series of Polyzoa, Brachiopoda, and Tunicata.
The Starfish Gallery, so called from one of the best-known types of the Echinoderma, contains an exhibition of the animals of this Class, as well as of the somewhat heterogeneous assemblage of creatures which are comprised under the popular name of Worms (Vermes). These animals possess greater attraction to students of Natural History than to the general public, and many, from their small size or the soft nature of their body, are not suitable for exhibition. Therefore no attempt has been made to show more than iva carefully selected number of the types of the larger groups. But the exhibition of very complete series, supplemented by models or figures, to illustrate the remarkable life history of some of these animals, also of specimens of the Worms which possess a special interest from their relation to man, render this Gallery particularly instructive to the student.
This guide has been prepared by Mr. E. A. Smith, Mr. F. J. Bell, and Mr. R. Kirkpatrick, who have special charge of the collections described.
In conclusion, thanks are due to Messrs. A. & C. Black, Messrs. F. Warne & Co., Messrs. Macmillan & Co., Crosby Lockwood & Son, and the Linnean Society for kindly allowing the use of clichés from illustrations in various works published by them.
| THE SHELL GALLERY. | ||
|---|---|---|
| PAGE | ||
| General Notes on Mollusca | 1 | |
| Systematic Arrangement | 7 | |
| Amphineura (Chitons, etc.) | 7 | |
| Gastropoda (Marine Univalves, Land-Snails, etc.) | 9 | |
| Lamellibranchia (Bivalves, Oysters, Cockles, etc.) | 29 | |
| Cephalopoda (Octopus, Cuttlefish, etc.) | 45 | |
| Index to principal Genera of Mollusca | 51 | |
| General Notes on Polyzoa | 54 | |
| Classification | 57 | |
| An account of the Brachiopoda | 74 | |
| Classification | 77 | |
| An account of the Tunicata | 83 | |
| Arrangement | 90 | |
| THE STARFISH GALLERY. | ||
| General account of the Echinoderma | 106 | |
| Crinoidea (Lily Stars, etc.) | 112 | |
| Asteroidea (Starfishes) | 113 | |
| Ophiuroidea (Brittle-Stars) | 114 | |
| Echinoidea (Sea-Urchins) | 114 | |
| Holothurioidea (Sea-Cucumbers) | 115 | |
| Vermes | Platyhelminthes (Tapeworms, etc.) | 116 |
| Nematodes (Round-worms, etc.) | 118 | |
| Annulata (Marine Worms, Earthworms, Leeches) | 122 | |
The Mollusca constitute one of the principal divisions of the Animal Kingdom, and include such animals as the Octopus, Cuttlefish, Snail, Slug, Whelk, Cockle, and Oyster.
They may be characterized as soft, cold-blooded animals, without distinctly marked external division into segments (as in Worms); their cerebral ganglia (the centre of the nervous system) lie above the commencement of the gullet, and are connected with the inferior ganglia by nerve-chords. Their heart consists of two or more chambers, and is situated on the dorsal side of the animal; it drives the blood into spaces between the various organs of the body. Only the Cephalopods possess internal cartilages, but all are without a bony internal skeleton; in the majority this is compensated by an external hardened shell which is formed (secreted) by the outer covering of the animal termed the mantle. |The shell.| The shell may consist of two parts (valves), as in the Oyster, or may be single, as in the Whelk and Limpet, or composed of a series of plates, as in the “Coat-of-mail” shells or Chitons: when well developed it is hardened by a rich deposit of carbonate of lime; but it may be gelatinous, as in Cymbulia, or altogether absent, as in Octopus; it may cover and protect the body, as in the Oyster, lie within the folds of the mantle, as in the Sea-hares (Aplysiidæ), or it may be quite internal, as in the horny “pen” of the Squid. It may be elongated, as in the Elephant Tooth-shell (Dentalium), cup-shaped, as in the Limpet, or spirally coiled, as in the Snail.
The mantle may form a free fold on either side of the body, as in the Bivalves, or it may become largely attached to the body-wall, 2as in the Snail or the Slug, and so give rise to an air-chamber, which, when its walls are richly supplied with blood, serves as a lung. The ventral surface of Molluscs is produced into the so-called “foot,” which may be very variously modified. The foot may be more or less hatchet-shaped, or curved and capable of serving as a leaping-organ, or sole-shaped and adapted for creeping; its margins may be produced into elongated processes, as the so-called arms of the Octopus, eight in number and provided with suckers, or of the Nautilus, where the arms are much more numerous, but shorter and without suckers. In the Cephalopods, also, another part of the foot may fold over from either side and form a median funnel, through which the water of respiration is driven outwards, causing the animal to move in the opposite direction—this part of the foot having, therefore, still the function of an organ of locomotion. By means of their muscular foot the Solenidæ, or Razor-shells, burrow in the sand, the Pond-Snails (Limnæidæ) crawl on aquatic plants and swim reversed on the surface of the water, the Limpet clings to the rock, and the Cockles and Trigonias take surprising leaps.
Upon the upper surface of the foot, in many Gastropods, a flat hard structure termed the operculum is situated, which, when the animal is retracted, partly or entirely closes the aperture of the shell. In some cases, as in the Turbos, it is very strong and of a stony nature, but in most instances it is horny. It is differently constructed in distinct families: it may be annular and multispiral, annular and paucispiral, subannular and ovate, or subannular and unguiculate. In the Nerites it is shelly, somewhat semicircular, closes the aperture of the shell, and is furnished with a stout projection on the straight edge, fitting like a hinge under the inner lip of the shell. A series of opercula is exhibited in side table-case C.
Thread-like processes on either side of the body, the so-called gill-filaments, often unite with those in front of and behind them, and so give rise to plates; these, when well developed, are best seen in the division to which the Oyster and the Mussel belong, and which, therefore, has been called the division of the plate-gilled Molluscs, or Lamellibranchia. Where the body is coiled or twisted on itself, as so often happens, the gills of one side may be altogether lost. Sometimes, as in Phyllirhoë, when the body is small and its wall thin, the gills (ctenidia) disappear altogether, and there is no special breathing-organ; in others the loss of the gill is compensated 3by the formation by the mantle of a lung; this is most often seen in the forms that live on land.
But these so-called gills may have other functions: in the Lamellibranchs, where there is no head and no special means by which the creature can obtain food, the delicate waving filaments or cilia with which they are covered cause currents in the surrounding water, by means of which minute organisms are brought to the mouth.
All Molluscs, except the Lamellibranchs, have a very remarkable structure developed in the floor of their mouth-cavities; on a basis of cartilage, which may be moved backwards and forwards by muscles, there is developed a horny plate, which may be of considerable length, and which has its upper surface covered with a number of more or less fine, flattened, or spiny outgrowths, which are known as teeth. This is the odontophore, tongue, radula, or lingual ribbon (see fig. 3).[1]
Eyes may be absent, as in nearly all the headless Lamellibranchs; but in other Molluscs they are generally present, and may be more or less well developed. An instructive series of stages is exhibited by the Cephalopoda. In Nautilus the eye remains an open pit; in Ommatostrephes two chambers appear, the anterior of which is bounded posteriorly by the lens, and is open to the exterior, so that sea-water enters it; in Sepia, finally, the anterior chamber becomes closed in front. We may observe that the eyes of all Cephalopods are at first pit-like, or pass through a stage which is permanent in Nautilus, one of the geologically oldest types.
Cephalic eyes have recently been noticed in Mytilus and Avicula.
Eyes of a more complicated structure, which are modified tentacles, are sometimes found on the edges of the mantle in Lamellibranchs (e.g. Pecten); these eyes resemble those of Vertebrates, and differ from those of most invertebrate animals in having the fibres of the optic nerve entering the distal and not the proximal ends of the retinal cells. Eyes of a similar construction are to be found on the back of the shellless Oncidium, and may be about one hundred in number.
Eyes of a remarkable character on the shells of some of the Chitons appear to be modified from tactile organs, and are innervated like the ordinary molluscan eye; they sometimes occur in enormous numbers, more than ten thousand being present on one animal (see wax model, Case 2).
In Cephalopods the ear, like the eye, is known to make its first 4appearance in the form of an open pit, the mouth of which gradually closes up, leaving only a narrow slit in communication with the exterior. It is probable that in many forms the so-called ear is an organ by means of which the mollusc becomes acquainted with changes in the surface over which it is passing; it is often found deeply imbedded in the substance of the foot, where it forms a closed vesicle.
There is no doubt that the carnivorous Gastropoda are gifted with a sense of smell, and throughout the series we observe patches of modified cells of the body-wall (the osphradium) which serve either as olfactory organs or as an apparatus for testing the nature of the water of respiration.
The sexes are distinct in the most highly organized Mollusca, but are united in the same individual in some of the lower forms, such as Land-Snails, the Opisthobranchia (including the Bubble-Shells, Sea-Slugs, &c.), and in some Bivalves. The reproduction of Mollusca is in all cases effected by means of eggs. In some instances the young are actually hatched within the oviduct of the parent, as in the Freshwater Snails (Vivipara); and apparently in most Bivalves the eggs are also retained within the valves until hatched.
The ova of many molluscs are deposited in masses enclosed in capsules. Some of them are very wonderful and complicated structures. Those of the Cuttles and their allies are clustered like grapes, each capsule containing but a single embryo; but in the Calamaries or Squids they form a radiating mass of elongated sacks, each containing from thirty to two hundred eggs, and it has been estimated that one of the spawn-clusters of the Common Squid (Loligo vulgaris) contains as many as 40,000 ova. Everybody knows the spawn-cases of the Common Whelk, found so abundantly on the sea-beach, consisting of a large number of yellowish capsules, heaped one upon another and forming an irregularly rounded mass. As many as five or six hundred capsules may be piled together in a single heap, each capsule containing several hundred eggs, of which perhaps only thirty or forty are hatched.
In other genera, as Tethys, Doris, Eolis, &c., the eggs are contained in a spirally rolled ribbon or strap-like structure; and some of the Naticæ build a somewhat similar capsule, composed of the eggs cemented together by sand and a gelatinous material, the whole forming two-thirds of a circle narrowed at the upper part.
Terrestrial Molluscs deposit, in comparison with their marine relations, but very few eggs. They are sometimes covered by a 5thin soft skin, but in certain groups, such as the large South-American Strophochili and the African Achatinæ, which include the largest of known land-molluscs, they are protected by a hardened calcareous shell, in some instances fully an inch in diameter. The freshwater forms (Limnæa and Physa) deposit from thirty to a hundred eggs enveloped in a gelatinous mass.
The number of eggs produced by some Bivalves is enormous. The Common Oyster is said to produce a million or more, and the American variety ten, or even sixty, times as many. Some of the River-Mussels are also very prolific, as many as two millions being sometimes the product of a single individual. A small series of the eggs of Land-Snails and of the egg-capsules of some marine Gastropods is exhibited in side table-case C at the side of the Gallery.
The ova of Mollusca may be gradually developed into the form of the parent, or there may be a free-swimming larva, which has a circlet of cilia near the anterior pole of its body (so-called “Veliger” larvæ), or there may be special larvæ, as in the case of the Freshwater Mussel, the “Glochidium,” as it is called, which has a toothed bivalve shell by which it can fix itself to fishes.
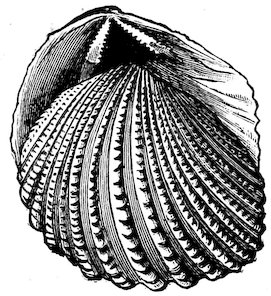
The limits of age of molluscs has been definitely ascertained in a few instances only. Most Land-Snails probably live about two years, although in confinement some have been kept alive for a much longer time. Some of the marine forms live for a considerable period, the Common Oyster not attaining full growth until about five years old, after which it may continue to live for many years. The Giant Clam, a specimen of which is placed in the upright cases near the entrance to the Gallery, must, one would think, have a very long existence, judging from the size and thickness of the shell. |Hibernation and torpidity.| All terrestrial molluscs hibernate in cold climates, hiding themselves away in the ground between roots and similar sheltered places. In tropical countries some assume a state of torpidity (æstivate) during the hottest and driest season of the year, closing up the aperture of their shells with a temporary lid or door (epiphragm), in order to resist the dryness of the atmosphere. Some of these “summer-sleepers” are endowed with a remarkable tenacity of life. An Australian Pond-Mussel has been known to live a year after being removed from the water; several Land-Snails have revived after a captivity of from two to five years, without any food whatever. One of the most remarkable instances of this kind occurred in the British Museum. A specimen of Helix desertorum, a common 6Desert-Snail from Egypt, was fixed to a tablet in March 1846, and in the same month of the year 1850 it was discovered to be alive. It must have come out of its shell in the interval, and finding it was unable to crawl away, had again retired within it, closing the aperture with a new epiphragm, but leaving traces of slime upon the tablet, which led to its immersion in water and subsequent revival, having passed a period of four years in a dry museum without the smallest particle of food. The actual specimen is here figured, Fig. 1.
Fig. 1.[2]
Helix desertorum.
(See black table-case 1.)
The economic uses of molluscs to man are manifold, and will be mentioned in the course of the description of the several families; but here may be the place to direct the attention of visitors to side table-cases B and D at the side of the room, containing some specimens of articles manufactured from shells, such as cameos, flowers, bracelets, brooches, &c.
Mollusca made their appearance on the globe at a very early epoch in the history of the development of animal life, a large number of fossil forms, such as Nautilus, Lituites, Orthoceras, &c., being found in the oldest Palæozoic formations. Probably all these belonged to the Tetrabranchia, of which one descendant only, the Pearly Nautilus, has survived to our period. Some Gastropods and Bivalves coexisted with those ancient Tetrabranchs; but these types abounded more in the later geological epochs, many Tertiary forms being undistinguishable from species which now exist.
The greater number of Mollusca are inhabitants of the sea, some passing their whole life at the surface hundreds or thousands of miles away from land; others at the bottom of the ocean at all depths, some having been dredged at five miles from the surface. Many are found in much shallower water, and a large number between tide-marks. Rivers and lakes furnish an immense variety of forms, and vast numbers live on land in all situations—on mountains, in valleys, forests, and deserts.
Molluscs are either animal or vegetable-feeders, the former preying principally upon other members of their own class.
7The following Table shows the systematic arrangement of the Mollusca adopted in the Shell Gallery:—
Class I.—Amphineura.
Class II.—Gastropoda.
Class III.—Scaphopoda. Tooth-shells (Dentalium).
Class IV.—Lamellibranchia. Bivalved Molluscs.
Class V.—Cephalopoda.
The Molluscs of this class are characterized by bilateral symmetry. The head and arms are situated at the opposite extremities of the elongated body, the gills, genital ducts and circulatory organs being paired and similar on both sides. The first order belonging to this division, the Polyplacophora, includes the “Coat-of-mail shells,” or “Sea-woodlice” (Chitonidæ). They have their back armed with eight shelly plates which overlap one another like tiles, and, like woodlice, have the power of rolling themselves into a ball. These plates are imbedded at the sides into the fleshy mantle, beneath which, on each side of the foot, are arranged the gills. A Chiton differs in many respects from other Mollusca. It has a shell like an Isopod Crustacean, a heart down the back like a sea-worm, symmetrical organs of reproduction on each side like the bivalves, a 8head and crawling foot like a true Limpet, and a posterior anal orifice. These several anatomical peculiarities at one time induced certain eminent authorities to hesitate in considering them molluscs; but now that the development from the egg has been investigated, their association with the Mollusca may be considered definitely settled.
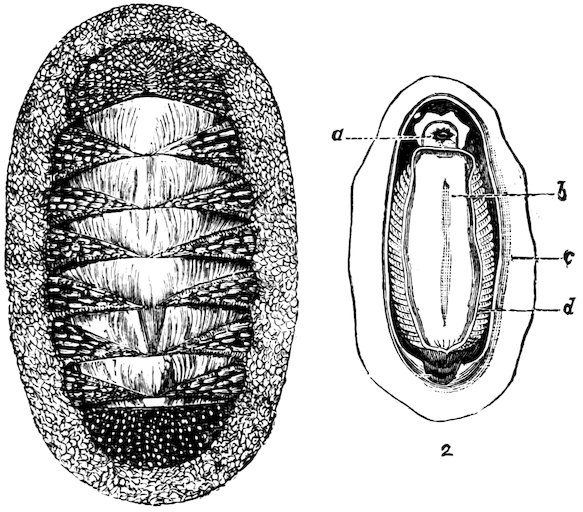
Fig. 2.
Coat-of-mail Shells, or Chitons.
1. Chiton squamosus (upper surface).
2. Chiton elegans (lower surface): a, mouth; b, foot; c, mantle; d, gills.
Chitons are found in all parts of the world, the finest inhabiting tropical countries. They live chiefly on rocks and under stones at low-water or at moderate depths; but a few forms have been discovered by the ‘Challenger’ Expedition at depths exceeding 2000 fathoms. The numerous sections of the group are principally distinguished by differences in the edges of the plates or valves which are inserted in the mantle, and in the different kinds of ornamentation upon the upper surface of the mantle-border. This, in some species, is quite smooth, in others covered with a dense mass of minute grains or scales, and in others armed with short prickly spines. In the giant Cryptochiton of Kamtschatka the plates are entirely covered over by the thick leathery granular mantle, and in another set, Cryptoplax, which consists of long slug-like animals, the plates are very small, and placed at intervals along the back.
About five hundred living species are known, and about one-fourth 9that number has been found fossil from the Silurian age downwards.
The second order of Amphineura, namely, the Aplacophora, comprises a few somewhat worm-like Molluscs which are devoid of a shell, but have instead the dorsal surface more or less studded with numerous minute calcareous spines or spicules. Neomenia, Proneomenia, Chætoderma are genera belonging to this order.
In contradistinction to the preceding class these Molluscs are asymmetrical, especially in respect of the gills and the spiral coiling of the viscera and most of the shells. They may be divided into two sections, Streptoneura and Euthyneura, distinguished by differences in the arrangement of the visceral nerve-loop.
The Molluscs of this section are bisexual and furnished with a shell, and generally with an operculum. The gills are in front of the heart and the visceral nerve-loop is twisted into a figure of 8. The section contains two groups or orders, Scutibranchia and Pectinibranchia.
The Scutibranchia have a free bipectinate gill, or the gill may be absent (Lepeta, Helicina), and generally exhibit traces of bilateral symmetry.
The Acmæidæ are called False Limpets, because, although the shells are identical with the true Limpets, the animals differ in having only a small gill on the left side of the neck, whilst the Patellæ have the gills greatly developed all round the sides of the foot. Both the true and the false Limpets are littoral and found on rocks between tide-marks. They have the power of excavating the surface to which they attach themselves, and adhere so firmly that it is easier to break the shell than detach the animal. The largest known Limpet (Patella (Ancistromesus) mexicana, case 7) inhabits the west coast of Central America, its shell having sometimes a length of 12 inches. The Limpets are vegetable-feeders and fond of seaweeds of various kinds, which they rasp with their remarkable spiny tongues. That of the common English Limpet (P. vulgata, Fig. 3) is longer than the shell itself, and armed with as many as 1920 glassy hooks in 160 rows of twelve teeth each. The Limpet is commonly used for bait in the sea-fishing off the Scottish coast, and vast quantities are consumed as food in some parts of Ireland. Some Limpets, such as P. compressa, P. mytilina, etc., are found on the stems of floating seaweeds, and have the shells usually thinner and smoother than the Rock-Limpets, which have to resist the fury of the breaking waves.
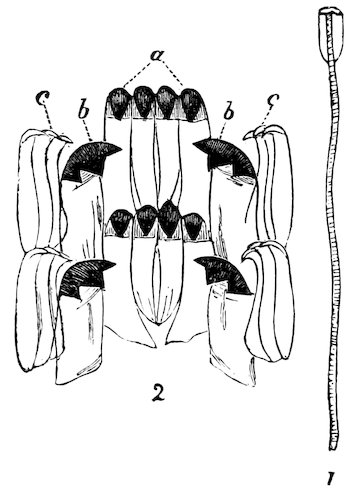
Fig. 3.
1. Radula of the Common British Rock-Limpet (Patella vulgata), natural size.
2. Two transverse series of teeth: a, median teeth; b, laterals; c, uncini or marginals.
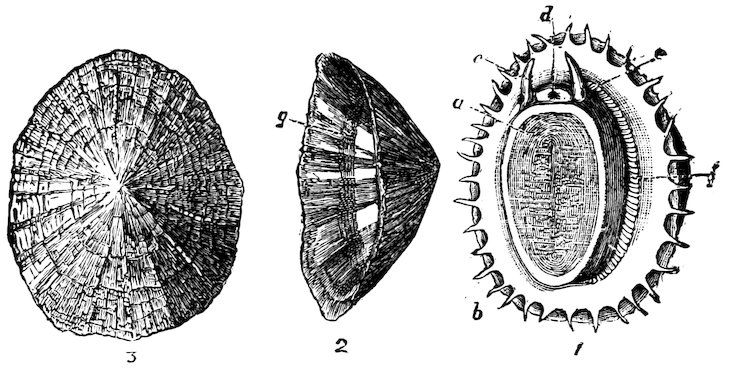
Fig. 4.
The Common Rock-Limpet (Patella vulgata). British.
1. Animal: a, foot; b, fringed mantle; c, tentacles; d, mouth; e, eyes; f, gills.
2. Side view of shell, showing the impression or scar of the attachment-muscle, g.
3. Upper surface of the shell.
11The “Keyhole Limpets” and “Slit Limpets” (Fissurellidæ) resemble in external shape ordinary Limpets, but are perforated at or near the apex, or more or less slit at the front margin. The hole or slit gives passage to a tubular fold of the mantle, through which the water apparently flows to the gills. The largest species are from California and South America, and others are found, but not abundantly, on most shores. The animal of the large Lucupina crenulata from California is eight or ten inches in length, and almost conceals the shell, and the shell of the South-African Pupillæa aperta is also all but hidden beneath the mantle of the animal.
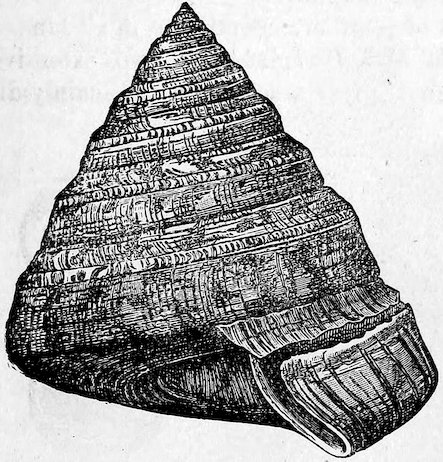
Fig. 5.[3]
Pleurotomaria adansoniana. Case 9.
⅓ natural size.
The Pleurotomariæ are extremely rare in recent times, only five species being known, whereas over a thousand fossil forms have 12been described. The specimens of P. adansoniana and P. beyrichi exhibited in case 9, are among the finest acquisitions to the shell collection of recent years.
The “Ear-shells” or “Ormers” (Haliotidæ) are found adhering to rocks in most parts of the world, with the exception of South America. They are lined with pearl, and many exhibit splendid colours and sculpture externally. Like the Limpets they hold on to the rocks with such tenacity that it is absolutely impossible to remove some of the larger species by force without injuring the shell. Boiling water or mustard and water poured over them will, however, soon compel them to relinquish their hold. The shell of Haliotis is pierced by a series of holes parallel with the left margin. Through such of them as are open the animal protrudes a slender filament or feeler, and the water also finds its way through them to the gills beneath.
The Single British species (H. tuberculata) is not actually found on the English coast, but common on rocks and stones at low-water in the Channel Islands. It is frequently eaten by the poor of those islands and the north of France; other species in New Zealand, China, Japan, West Africa, and elsewhere, constitute a common article of diet among the natives. Haliotis-shells are largely used in the manufacture of pearl ornaments, and in all kinds of inlaid work.
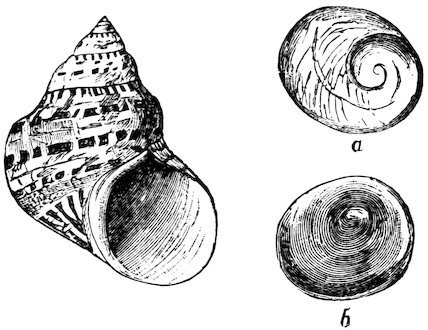
Fig. 6.
Top-shell (Turbo petholatus). (From the Indo-Pacific Ocean.)
a. Inner surface of operculum. b. Exterior of ditto.
The Trochidæ, and Turbinidæ are two extensive families, the animals of which are very much alike, and mainly distinguished by the operculum, which in the former is horny, and shelly in the latter. The shells of these families are beautifully pearly within, 13and the external shelly coat is generally brightly coloured and highly ornamented. Several very pretty species are found on our own shores. The opercula of Turbo petholatus (Fig. 6), from the Indian and Pacific Oceans, are frequently mounted in gold and silver as scarf-pins, ear-rings, &c.
The Nerites are mostly found in tropical countries, and, like the Winkles, are very strongly made, to resist the force of the breaking waves. The Neritinas are partly found in the sea, and partly in fresh water, and are less solid shells. The third section of Neritidæ, the Septariæ, are shaped very much like Limpets, except that the apex is at one end instead of central. They are, however, very different animals, and furnished with a shelly operculum imbedded in the foot.
In most cases the molluscs of this order have an attached monopectinate gill and a single osphradium. A few are fresh or brackish water forms, but the majority are marine.
The “River-Snails” (Viviparidæ) might be termed freshwater Periwinkles, as the animals of both are very similar. The true Viviparæ are viviparous. They are rather sluggish, and found at the bottom of ponds and rivers feeding on decaying animal and vegetable matter.
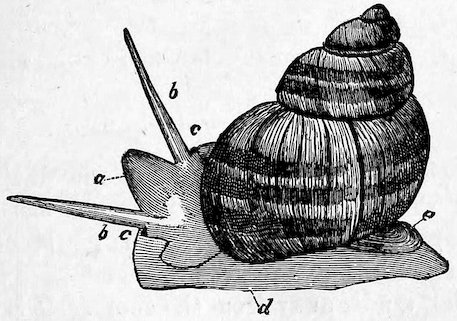
Fig. 7.
The Common British River-Snail (Vivipara vivipara).
a, head; b, tentacles; c, eyes; d, foot; e, operculum.
The Cyclophoridæ are land-shells, which, however, cannot properly be considered true lung-breathers like ordinary snails. They have not the closed lung-chamber of the Pulmonates, their eyes are 14placed at the base of the tentacles instead of at their tips, they have a long proboscis armed with a different rasping tongue (radula), a spiral operculum, and the sexes are distinct, whereas the true Snails are hermaphrodite.
The operculated air-breathers have been divided into many sections, chiefly on account of differences in the apertures of the shells and in the opercula. They most abound in hot countries, but a few species are met with in temperate regions.
The “Apple-Snails” (Ampullariidæ) live in the rivers and marshes of tropical regions, and, although represented by a large number of species, exhibit comparatively slight variations in form and colour. The animal has both a pectinated gill and a lung cavity, being thus enabled to breathe either water or air.
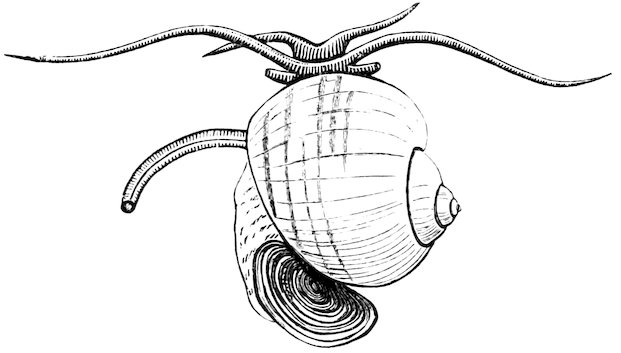
Fig. 8.
Ampullaria canaliculata.
The “Periwinkles” (Littorinidæ) are found almost on every known shore; they feed upon all kinds of marine vegetation. Some species are met with at low-water mark, others on rocks almost beyond the reach of the sea, and some have been discovered inland nearly half a mile away from the shore. It is calculated that 1900 tons of the “Common Periwinkle” (Littorina littorea), of the value of £15,000, are annually consumed in London alone.
The family of Calyptræidæ includes the “Slipper-Limpets” (Crepidula) and the “Cup-and-saucer Limpets” (Crucibulum). Although furnished with a foot, they rarely crawl about, but remain attached to rocks, stones, or other shells, sometimes forming a shelly plate under the foot by which they become fixed to the spot where they have taken up their abode.
The “Cowry-shells” (Cypræidæ) are remarkable for their varied 15markings and splendid polish, which is produced and preserved by two flaps of the mantle, one on each side, which fold over the back, a line down the centre of which usually marks where the flaps meet. The animals are even more brilliantly coloured than the shells. They have no operculum, but a large foot, which they can withdraw entirely within their shell, although the aperture is usually very narrow. Cowries, as is well known, are sold as ornaments; and a small yellow species, “the money-cowry” (C. moneta), which is very common in the Indian and Pacific Oceans, passes current as coin among the negro tribes of certain parts of Africa. The specimen of Cypræa leucodon figured on p. 16 is extremely valuable and supposed to be the only one hitherto discovered. The “orange cowry” (Cypræa aurora) is worn by chiefs in the Friendly Islands, and is considered the highest order of dignity. Only one small species, Trivia europæa, is found on the British coast, and about 100 fossil forms have been discovered in the Chalk.
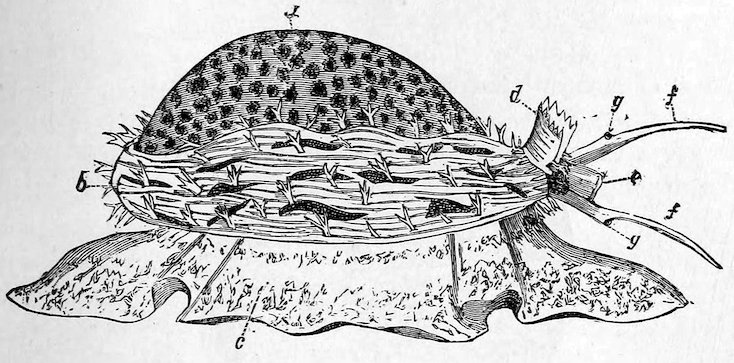
Fig. 9.
The Tiger Cowry (Cypræa tigris). (From the Indo-Pacific Ocean.)
a, the shell; b, the mantle; c, foot; d, siphon; e, proboscis; f, tentacles; g, eyes.
Of the Ovulidæ, the most curious is the “Weaver’s-shuttle” (Radius volva), in which the shell is peculiarly beaked at both ends. It is found living on barked corals (Gorgoniidæ), and some of the smaller species exhibit differences of coloration, resembling the tints of the Gorgonias upon which they are found.
The Naticas are mostly blind, and have a very large foot, suitable for burrowing in the sand when in quest of bivalves. They are very 16voracious. This is one of the groups of shells that have continued to exist from Palæozoic times.
The “Violet Snails” (Ianthinidæ) are found floating about in every ocean, excepting in cold regions, with the spire of the shell downwards, and the bottom, being more exposed to the action of light, is more deeply tinted than the upper part. They feed upon Jelly-fish, and construct a gelatinous raft, filled with air-bubbles, beneath which the females attach their eggs.
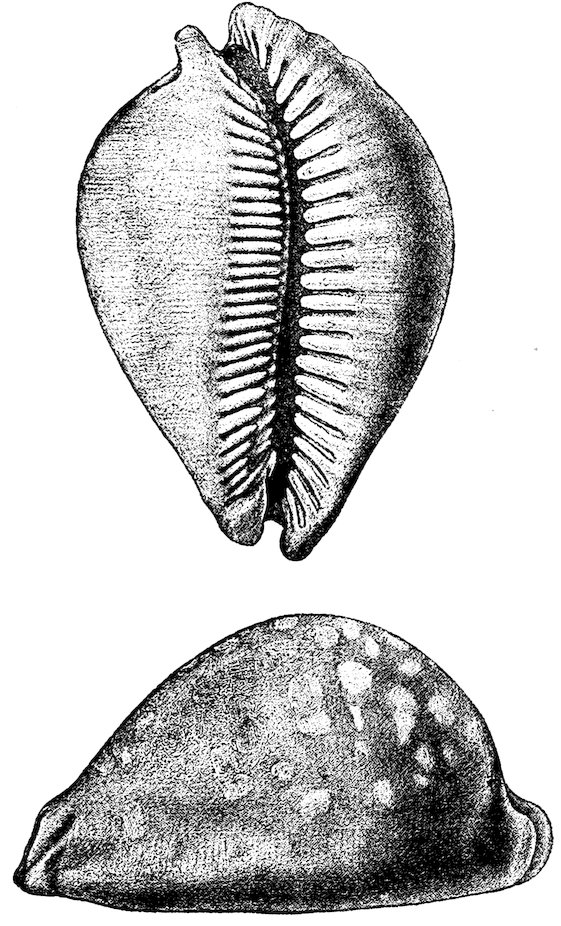
Fig. 10.
Cypræa leucodon. Case 32.
17The Melaniidæ are freshwater Snails which abound in most tropical and subtropical countries; about 1000 species are known. They are mostly of dark colours, and are fond of muddy places.
The Cerithiidæ are chiefly marine forms, some, however, entering brackish water. About five hundred fossil species have been described, some of them gigantic in comparison with any now living, of which more than two hundred are known.
The Scala scalaris was formerly considered a great rarity, as much as £40 having been given for a single specimen, which might now be purchased for as many pence.
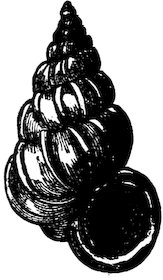
Fig. 11.
Scala scalaris.
Case 44.
The “Worm-shells” (Vermetidæ) are a very peculiar family. Their shells can scarcely be distinguished from the shelly tubes which are formed by certain species of marine worms, Serpula, &c. They are free and spiral in early life, but afterwards become distorted and generally attached to rocks, stones, &c. A foot for walking purposes therefore would be of no use; consequently it is more or less obsolete, serving only as a support to the operculum.
The “Screw-shells” (Turritellidæ) have elongate tapering shells; about 100 recent and 200 fossil species are known. One species only (Turritella communis) is now found living on the British coasts.
The Xenophoridæ have the singular habit of cementing to the exterior of their shell, stones, pieces of coral, and fragments of other shells; hence they have been called “Carrier-shells,” and, according to the kind of material chosen, have been named “Conchologists” and “Mineralogists.” Beyond acting as a disguise, and consequently as a protection, there does not appear to be any special utility in thus adding to the weight of their own shells. The animals do not glide like most other molluscs, but scramble along like the Strombs, the form of their foot being small, divided into a front, expanded, and a hind, tapering portion admirably adapted to the nature of the ground on which they live, which usually consists of broken and dead shells.
The “Wing-shells” (Strombidæ) are the largest of the Gastropods with a proboscis or non-retractile snout. They do not crawl like most other Gastropods, but progress by a sort of hopping movement. They act as scavengers, feeding on decomposing animal matter.
The Strombus gigas, or “Fountain-shell,” occurs in great numbers 18in the West Indies, and is a very heavy solid shell. It is a favourite ornament for rockwork and fountains in gardens, and, like the Helmet-shells, is used for cameo-carving. It is also employed in the manufacture of porcelain, as many as 300,000 having been imported into Liverpool in one year for that purpose.
The Scorpion-shells, or “Spider-claws,” as they are sometimes called (Pterocera), possess singular claw-like projections, which are developed on the outer lip of the shells.
The “Trumpet-shells” (Lotoriidæ) have varices or strengthening ribs at intervals, like the Murices; the largest species, Lotorium variegatum, is used by South-Sea Islanders as a horn or trumpet. A hole is made in the upper part of the spire to blow through, and the sound produced can be modulated or varied by inserting the hand in the aperture or mouth of the shell.
The “Helmet-shells” (Cassididæ) are used for cameo-carving; they consist of differently coloured layers, so that the ground-colour of the carving is of a different tint from the subject engraved. The most artistic shell-cameos are produced in Italy, whence the art has been introduced into France and England. The Cassis madagascariensis (Fig. 12 on p. 19) is in special request by shell-carvers on account of the strong contrast of the white upper layer with the dark ground beneath. Extinct forms of Cassis are found fossil in Tertiary formations, but none of them equal in size the largest living species.
The “Tun-shells” (Doliidæ) are remarkable for the globoseness of the shells, which are covered with very regular revolving ribs.
The Fasciolariidæ contains two of the largest living Gastropods: Megalatractus aruanus, from North and West Australia, and Fasciolaria gigantea, which is found off the coast of South Carolina, and attains at times a length of two feet.
The Mitras (Mitridæ) are great favourites with shell-collectors, on account of their beautiful colours and varied sculpture. There are about 600 living species already known, and between one and two hundred have been found in a fossil state. Shells of this group, like the Fasciolariæ, are distinguished by a few plaits or folds on the inner side of the aperture (the columella). Mitras are almost exclusively found in tropical or subtropical regions, the majority being met with either at low-water mark or in comparatively shallow water.
The family of Buccinidæ also contains a very large and various assemblage of forms. Among them may be mentioned the Whelks (Buccinum). (See Fig. 13 on p. 19.)

Fig. 12.
Cassis madagascariensis, with cameo engraved upon it.
Side table-case B.
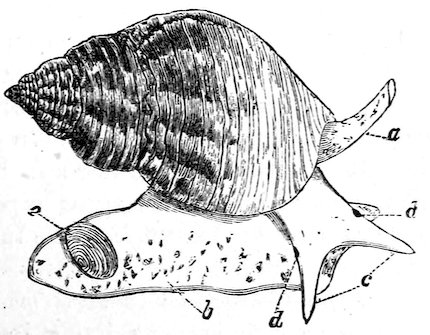
Fig. 13.
The Common Whelk (Buccinum undatum).
a, siphon; b, foot; c, tentacles; d, eyes; e, operculum.
20The family of Muricidæ, or “Rock-shells,” is another extensive group, containing many very handsome and peculiar forms. The animals of this family have a long proboscis, at the end of which is the spiny tongue (radula), and which is retractile within the body. The true Murices produce at intervals ribs or varices, which in some species are ornamented with long spines or foliations, and which indicate periods of growth, but of what duration we do not know. They are all carnivorous, feeding chiefly on other Mollusca, boring through the shells of bivalves with their spiny tongue, and slowly devouring the unfortunate inhabitant piecemeal. From certain species of Murex (M. brandaris, &c.) found in the Mediterranean, the ancients manufactured the celebrated Tyrian purple dye.
The “Purples” (Purpura) are found between tide-marks all over the world. Magilus, belonging to the family Coralliophilidæ (Case 77), is found among coral-reefs in tropical seas, and has the remarkable habit of lengthening the aperture of its shell into an elongate tube, in order to keep pace with the growth of the coral, and to prevent its being overgrown and killed.
The “Volutes” (Volutidæ) are a group of shells also much sought after by shell-collectors. Some of these attain to a very large size, the animals inhabiting them being enormous. The Boat-shells (Cymba) and Melons (Cymbium) are ovo-viviparous, the young being carried about by the parent until they are an inch in length. Volutes are found chiefly in the warmer parts of the Atlantic and Indo-Pacific Oceans, and occur in the greatest variety on the coasts of Australia.
The Olives (Olividæ) are common in most tropical seas, and are remarkable for their beautiful polish and various patterns of colouring. In structure and form they are very similar to each other. They burrow in sand in quest of bivalves for food, and some species are said to have the power of swimming by expanding the lobes of the foot.
The Harps (Harpidæ) form a small well-marked group, of which probably nearly all the existing species have been discovered. The animals inhabiting these beautiful shells are also brightly coloured. They have the remarkable power of casting off a portion of the foot when disturbed. The species are known from the Indo-Pacific Ocean, the west coast of Central America, and West Africa.
The next family, the “Slit-lips” (Pleurotomatidæ), consists of very numerous species, over a thousand living forms having been discovered, and almost as many fossil species from Cretaceous and 21Tertiary strata have been described. The typical forms are characterized by a slit in the outer side (lip) of the aperture. Species of Pleurotoma are found in every sea, although most abundant in the tropics, and, although so numerous in species, the number of specimens is small in comparison with some other genera.
The “Auger-shells” (Terebridæ), like the Cones, present a great similarity in form, but, unlike them, have a great diversity of “sculpture” or external ornamentation. They are all elongate shells, with a deep notch at the base of the aperture. Owing to the length and comparative solidity of the shells, the animals of many of the species do not carry their shelly structures on their backs, like most other species, but drag them along the sandy sea-bottom.

Fig. 14.
The “Glory-of-the-Sea” Cone (Conus gloria-maris). Case 94.
(From the Philippine Islands.)
The Conidæ, or Cones, form one of the most beautiful portions of the collection of Shells. This family, of which between 400 and 500 22distinct kinds are known, is a great favourite with collectors on account of the brilliant colours and various patterns of the shells. Some, owing to their beauty and rarity, have been sold at very high prices, as much as £50 having been paid for a single shell. The Cones are found in all tropical seas, but are rare in cold or temperate latitudes. None are met with on our own shores, one species alone being known from the Mediterranean. They occur fossil in the Chalk and Tertiary strata. These animals are all carnivorous, and live usually in shallow water among rocks and coral-reefs. Some of them are said to bite when handled, and to be dangerously poisonous, the bite in some instances having been all but fatal.
The Atlantidæ, Pterotracheidæ, and Carinariidæ, at various times recognized as forming a distinct sub-class or order of Gastropoda, under the name of Heteropoda or Nucleobranchiata, are now regarded as families of aberrant Gastropods organised for swimming in the open sea. The Atlantas are found in great numbers in warm latitudes, and are provided with a glassy, thin, flat, spiral shell, not unlike a keeled Ammonite. The glassy shell of the Carinaria is one of the most beautiful structures of any mollusc, and at one time was such a rarity that £100 are said to have been given for a single specimen, which at the present time is perhaps worth only from five to ten pounds. Species of Carinaria are found in the Mediterranean and warmer parts of the Atlantic and Indian Oceans. The animal is large, semitransparent, and elongate, with a compressed fin-like foot which projects from the body, and is used in swimming. The gills are placed towards the hinder part of the back and covered by the shell. They feed on jelly-fish of various kinds, and probably on other soft animals.
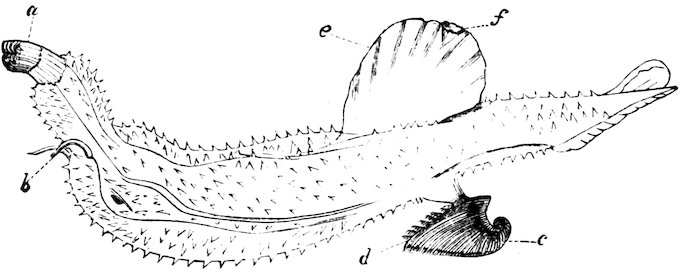
Fig. 15.
Glassy Nautilus (Carinaria lamarcki).
a, proboscis; b, tentacles; c, shell; d, gills; e, foot; f, sucker.
The Gastropods belonging to this sub-class have the visceral nerve-loop straight and not twisted as in the Streptoneura. All the Euthyneura are hermaphrodite, and their radula is generally composed of numerous similar denticles on each side of a median tooth. Scarcely any of these forms are provided with an operculum in the adult state. The Euthyneura may be divided into two orders, Opisthobranchia and Pulmonata.
All the Molluscs of this order are marine, some (Tectibranchia) breathing by means of the ordinary Gastropod ctenidium, which is generally behind the heart, whereas others (Nudibranchia) have developed a different type of respiratory organs.
The Opisthobranchia include the “Pteropods” formerly considered as a distinct class, the “Bubble-shells” (Bullidæ), the “Sea-Hares” (Aplysiidæ), the “Umbrella-shells” (Umbraculidæ), the Nudibranchs and some others.
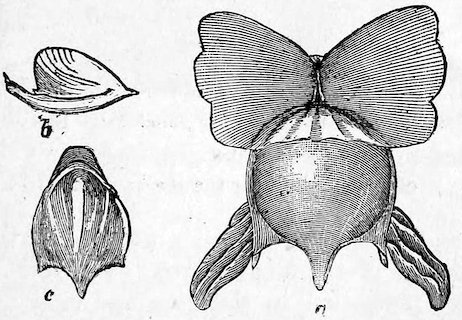
Fig. 16.
Shell-bearing Pteropod (Cavolina tridentata). Case 96.
a. Shell and animal. b. Side view of shell. c. Dorsal view of shell.
The Pteropods[4] are sometimes called Sea-butterflies, and are organized for swimming freely in the ocean. They have a pair of fins developed from the sides of the mouth or neck, which perform a flapping movement during progression. Some Pteropods (Thecosomata) 24are provided with small glassy shells; others (Gymnosomota) are naked. They exist in countless millions in some parts of the ocean, discolouring the water for miles. They constitute the principal food of the Baleen Whales.
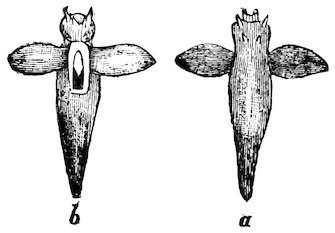
Fig. 17.
Shell-less Pteropod (Clione limacina).
a. Dorsal view. b. Ventral aspect.
About a hundred species are known.
The Sea-Hares, so called on account of a slight resemblance to a crouching hare and not for their nimbleness of foot, are found in most parts of the world, in pools at low water. At the hinder part of the back two flaps of the mantle partly conceal a thin horny shell which serves as a protection to the gills and vital organs beneath. When molested, these animals discharge a large quantity of a purple fluid, discolouring the surrounding water for a distance of more than a yard.

Fig. 18.
Sea-Hare (Tethys (Aplysia) punctata). British.
a, labial tentacles; b, upper tentacles or rhinophores; c, siphonal fold of the mantle near the shell; d, eye.
The shell of Umbraculum is shaped very like that useful article, an umbrella, of the Chinese pattern. The animal is very large, having its breathing-organs on the right side below the shell.
The Nudibranchs or Naked-gilled Molluscs comprise some of the 25most beautiful and strange forms. They are unprovided with shells except in the earliest stages of their existence, when they dwell in a minute nautiloid shell, furnished with an operculum, both of which are subsequently cast off. Unfortunately the colours of these beautiful creatures cannot be preserved after death, and therefore a small series of glass models is exhibited, which will give some idea of their great variety in form and colouring. They are found in most parts of the world, chiefly in shallow water, but a few species live upon floating seaweed in the open sea. Over a hundred species exist on the British coast, the majority of which are, however, very small. They are chiefly carnivorous, feeding on other molluscs, sea-anemones, &c.
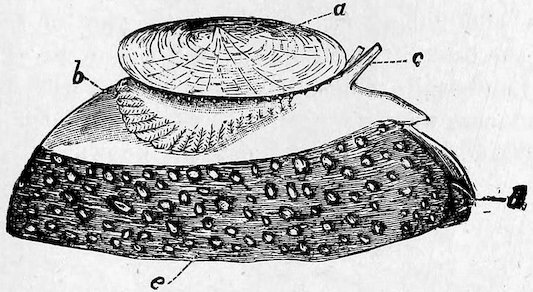
Fig. 19.
The Umbrella-shell (Umbraculum mediterraneum).
a, shell; b, gills; c, tentacles; d, mouth; e, foot.
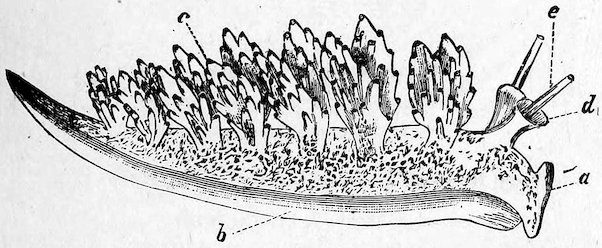
Fig. 20.
Naked-gilled Mollusc, or Nudibranch (Doto coronata).
a, head; b, foot; c, gills; d, tentacle-sheath; e, tentacle.
The Pulmonata are furnished with a lung cavity in place of the ordinary gill of other Gastropods, and may be termed true air-breathers. 26Most of them are provided with shells, and, with the exception of the Amphibolidæ never possess an operculum. They are divisible into two groups or sub-orders, Basommatophora and Stylommatophora, characterized by a difference in the position of the eyes. The Basommatophora, including the Auriculidæ, Amphibolidæ, Siphonariidæ, and Limnæidæ, have a single pair of non-retractile tentacles, at the base of which are situated the eyes. The Stylommatophora (Land-snails, Slugs, &c.), are provided with two pairs of retractile tentacles, with the eyes at the summit of the upper pair. Over ten thousand species of Pulmonata are known.
The first group of the aquatic air-breathers, the Auriculidæ, chiefly inhabit salt or brackish water. The largest forms are tropical and found at the mouths of rivers, among the roots and stems of mangrove-trees, or in damp woods near the sea.
The “Limpet-Snails” (Siphonariidæ) seem at first sight to be out of place among the Snails and Slugs, and more nearly allied to the Rock-Limpets; but the character of the tongue (radula) and the closed respiratory cavity indicate a close relationship with the present group.
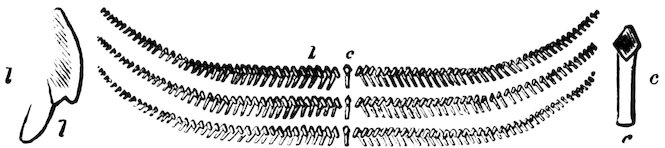
Fig. 21.
Three Rows of Teeth of the Radula of Siphonaria.
c, central; l, lateral teeth.
The shells of Siphonaria may be known from Limpets by a slight bulging on one side, caused by a radiating groove which interrupts the muscle of attachment. They are marine, and are found on rocks between tide-marks, chiefly in tropical countries.
The Limnæidæ are only found in fresh water. Most of them occasionally rise to the surface to breathe, where they glide along foot uppermost, at times suspending themselves by a glutinous thread, after the fashion of a spider. All countries appear to have their peculiar species.
27The freshwater Limpets (Ancylus) live attached to stones and leaves of plants, and have not the habit of floating, but, like the rest of the Limnæidæ, feed on freshwater algæ, confervæ, and decayed vegetable matter.
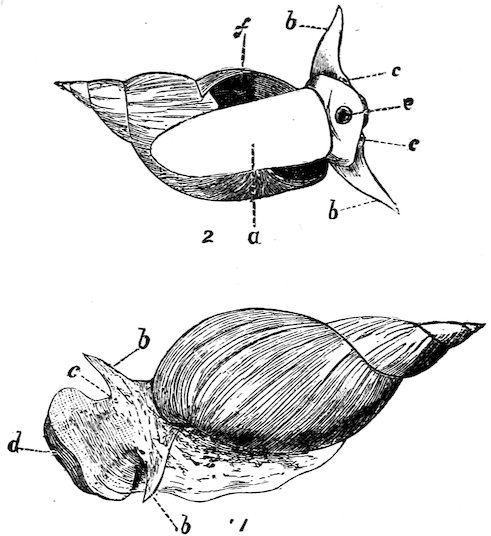
Fig. 22.
British Pond-Snail (Limnæa stagnalis).
1. Upper view: a, foot; b, tentacles; c, eye; d, muzzle.
2. Lower view: letters a, b, c as above; e, mouth; f, respiratory orifice.
True Snails (Helicidæ, etc.) have a distinct head furnished with eyes, tentacles, cutting upper jaws, and rasping teeth, and all are protected by a spiral shell. They are almost exclusively vegetable-feeders, subsisting chiefly on leaves. The sexes are not distinct. Many of the species are beautiful objects on account of the brilliancy of their colouration, and some are remarkable for the variation they exhibit in this respect. Species of Helicidæ are found in nearly every part of the world and in all situations, from sea-level to an altitude of 12,000 feet. They are fond of moisture, and in hot and dry weather retire within their shells, remaining torpid until the return of dew and rain. Helix pomatia (Case 119), which is found on the chalk in the south of England and on the Continent, is commonly eaten in Austria, France, and Belgium.
28The eggs of Land-Snails vary in texture, size, and in numbers: they are usually white, but in some instances yellow and pale green. Those of some of the large South-American forms are as hard as that of a hen, and more than an inch in length (Case 120).
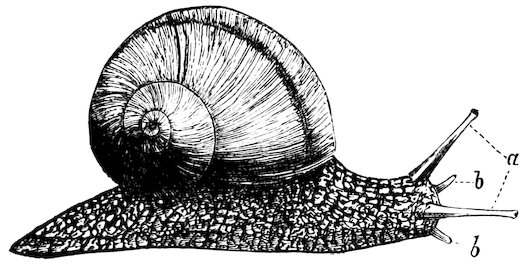
Fig. 23.
British Land-Snail (Helix pomatia).
a, eye-bearing tentacles (“horns”); b, lower or smaller tentacles.
Slugs (Cases 106,107) are very like Snails without external shells; most of them, however, possess a small internal shelly plate, or a few calcareous granules hidden beneath the skin of the back. Some have a large slime-pore at the end of the foot, and others are slightly phosphorescent. Like the Snails, they are fond of damp localities, and at times become great pests to farmers in devouring the young shoots of the growing corn. Testacella, which is found in this country, differs from the Slugs in having an external shell at the tail-end of the foot. It is not slimy, and lives under ground, feeding upon earthworms.
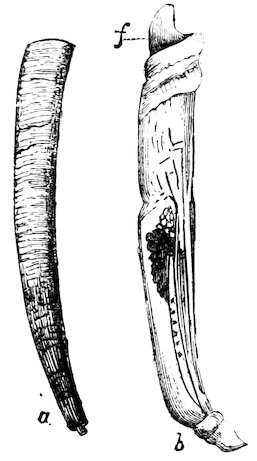
Fig. 24.
British Tooth-shell.
(Dentalium tarentinum).
a. The shell. b. The animal, removed from its shell; f. the foot.
The “Tooth-shells” (Dentaliidæ) form a distinct group, the shells of which are very unlike those of any other mollusc, but closely resembling the shelly tubes constructed by certain kinds of marine worms. The Dentalia 29have neither eyes nor tentacles, or a distinct head like Gastropods; their organs of circulation and respiration are of a rudimentary kind, and they have no heart. The sexes are separate. Their foot is adapted for burrowing in sand, in which they live and obtain their food, which consists of Foraminifera and minute Bivalves. One species, Dentalium pretiosum, found on the shores of North-West America, was until recently used as money by the Indians.
The Molluscs belonging to this Class have neither head, nor cephalic eyes, nor jaws or tongue like those of the other Classes, and are enclosed in a shell which consists of two plates or valves held together on one side of the margin by a horny, elastic substance, called the “ligament.” Bivalves do not creep about in search of food, but find their means of existence in the shape of minute particles, both animal and vegetable, which happen to be contained in the water which they breathe. Some, however, are capable of locomotion by means of a well-developed foot, and a few swim through the water by alternately opening and shutting their valves. The body is enclosed within two lobes of the mantle which line the interior of the valves, and which at their base are firmly attached to the shell, producing on the shell a scar or impression called the “pallial line.” The gills are lamellar or leaf-like, and placed on each side of the body. Each gill is called a ctenidium, and consists of an axis which is partly attached to the body of the Mollusc. This axis generally gives off two plates consisting of hollow filaments which are parallel with one another, directed downwards towards the ventral side, and in most cases long and refolded upon themselves, so that each plate becomes in reality a double lamella. In a few instances, however, the filaments are simple and not reflected. They are connected with one another by microscopic cilia, sometimes by vascular junctions, and the dependent and reflected portions (lamellæ) of each filament may be connected by “interlamellar vascular junctions.” The mouth is merely an oval aperture at the anterior end of the body, and generally furnished on each side with 30soft thin flaps, or labial palps, which have the function of conveying the food to the mouth. The mantle secretes the substance out of which the shell is formed. The two valves are always in contact at the hinge, which is generally formed by small interlocking projections or hinge-teeth, and they are closed by large adductor muscles, which are attached to impressions in the interior of the shell. When these muscles cease to act, as after death, the valves of the shell open in consequence of the elasticity of the ligament on the dorsal margin. The majority of species have two principal adductors, one at each end, like the Venus-shells, Cockles, Razor-shells, &c.; but in Oysters, Scallops, and a few others, there is but a single central muscle. All Bivalves are aquatic, and the majority marine. They are found burrowing in sand or attached to rocks. Some perforate stones and corals, others wood, and a few construct a sort of nest of fragments of shells, stones, &c.
Many schemes of classification have from time to time been propounded, based upon the presence or absence of respiratory siphons, the number and position of the adductor shell-muscles, the character of the shell-hinge, &c. The most recent arrangement is founded principally upon the structure of the gills. The value of such a classification has yet to be fully tested. Mr. Paul Pilseneer has suggested five orders of Lamellibranchs: Protobranchia, Filibranchia, Pseudolamellibranchia, Eulamellibranchia, Septibranchia.
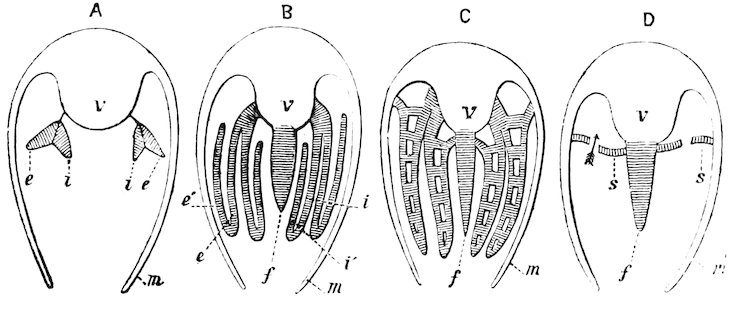
Fig. 25.
(From the ‘Cambridge Natural History.’ Messrs. Macmillan & Co.)
A. Protobranchia. B. Filibranchia. C. Eulamellibranchia. D. Septibranchia.
m. Mantle, v. Body. f′. Foot. e. Outer gill-lamella; i. Inner gill-lamella; e′. Reflected portion of outer lamella; i′. Reflected portion of inner lamella; s. Septum-like gill.
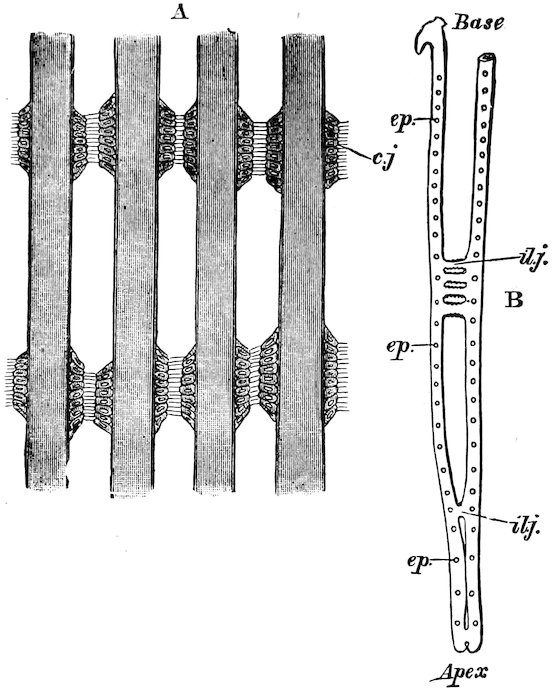
Fig. 26.
Gill of Mytilus edulis.[7]
A. Part of four filaments showing ciliated interfilamentar junctions (cj).
B. Diagram of a single filament showing the two lamellæ connected at intervals by interlamellar junctions (ilj) and the position of the interfilamentar ciliated junctions (ep).
In this order the filaments of the gills are not reflected, but arranged in two divergent rows, the foot being expanded and flattened beneath with crenulated margins and with the byssal gland very slightly developed. The Nuculidæ and Solenomyidæ are the only families belonging to this order. The shells of the former are remarkable for the numerous fine interlocking hinge-teeth, and those of the latter on account of the strong fringed periostracum.
In this group the gills are smooth, with the filaments directed downwards, reflected, and connected one with another by interfilamentar ciliated junctions, but the lamellæ are not connected. The foot is usually provided with a well-developed byssal gland. Anomia, Arca, Trigonia, and Mytilus belong to this order.
The family of Anomiidæ contains a number of more or less pearly shells remarkable for a deep notch or hole in the lower or flat valve through which a shelly plug passes, by means of which the animal attaches itself to other shells, stones, &c. Anomia ænigmatica is found adhering to leaves in mangrove-swamps.
The Placunidæ, sometimes called Window-shells and Saddle-Oysters, are very flat pearly shells with a remarkable hinge, which consists of two long divergent teeth, like a ⋀, to which the ligament is attached. The species are few in number, and inhabit sandy shores of India, China, and North Australia.
The Arcidæ are a family of strong ponderous shells varying much in form and sculpture. The animals have a longish pointed foot, deeply grooved along the bottom, no labial palpi, and free margins to the mantle, which are not prolonged into breathing-siphons. Many of the Arks often anchor themselves by means of a strong byssus. The shells of this family are usually radiately ridged; and the hinge is composed of a number of teeth arranged along the hinge-line, which is generally straight. Arca tortuosa, from China, has the valves curiously twisted. The section Barbatia is remarkable for the coarse fibrous character of the epidermis; Scapharca for its unequal valves; and Cucullæa, from the Indian Ocean, for the elevated ridge bounding the posterior muscular impression. Glycymeris (better known as Pectunculus) has the hinge-teeth arranged in an arched series, and the shells are more regular in growth than in many other forms of Arcidæ.
The Trigoniidæ are one of those families which have all but disappeared during our period. Only three or four living species are known, whilst more than a hundred fossil forms have been described from the Jurassic and Cretaceous formations. Australia, where some of the oldest types of animal life persist, furnishes also the existing species of Trigonia (Fig. 27). The animals have a long, sharply-bent, pointed foot like the Cockles, with which they make surprising leaps. The shells are beautifully pearly within, and ribbed and noduled exteriorly.
The Mytilidæ, or Mussels, are too well known to need description. The small foot, which is brown in the common species, is not much used in creeping about, but has the power of spinning a byssus or bundle of tough threads, by means of which the animals attach themselves to rocks and one another, forming colonies of vast numbers. Mussels have always been much eaten in this and other maritime countries, and large quantities are brought to the London market from the Dutch coast. At times they are unwholesome; but all the exact causes of this are not known. Mussels seem to be found on every shore, and some of the species are very widely distributed—the common edible Mussel, M. edulis, being found on every European coast, on the shores of North and South America, in the Arctic and Antarctic Oceans, and probably on the coasts of Australia.
One group of Mussels (Lithodomus, Case 144) burrow in rocks and other shells, forming holes just large enough to contain their shells. L. dactylus is sold as an article of diet on the shores of the Mediterranean.
The gills in this order are plicate, and the two lamellæ of each plate are furnished with conjunctive or vascular interlamellar junctions, and the filaments are connected by interfilamentar ciliated discs in some cases, in others by vascular concrescence. The mantle-margins are separated all round, and the foot is either small or wanting. Only a single adductor muscle is generally present. The Pearl-oysters, true Oysters, and Scallops are the forms which constitute this order.
The large family of Aviculidæ includes the “Wing-shells” (Avicula), the “Pearl” and “Hammer Oysters” (Meleagrina and Malleus), and Pinna (Fig. 28). Some species of Pinna attain to a length of two feet. They are found imbedded in the sand with the narrow pointed end downwards. They form a large silky byssus, which can be woven or knitted into gloves, socks, etc. (see side table—case B). The “Hammer-Oyster” (Malleus) is so called from its rude 34resemblance to a hammer. The “Pearl-Oysters” (Meleagrina margaritifera, Fig. 29) possess rather heavy strong shells, lined with very thick layers of “mother-o’-pearl.” Hundreds of tons of these shells are annually collected at the great pearl-fisheries of North and West Australia, and imported into Europe. The pearl-oyster of Ceylon (M. fucata, Case 146) is a smaller species, and collected more for the pearls than the shells. The round pearls, which are valued so highly, are either excrescences of the pearly layer or are found loose in the fleshy parts of the animal. Some small foreign body which has accidentally penetrated under the mantle and irritates the animal is covered with successive concentric layers of nacre, thus attaining sometimes, but rarely, the size of a small filbert. The nacre is generally of the well-known pearly-white colour, very rarely dark, and occasionally almost black. The action of the animal in secreting successive layers of nacre over any foreign body which intrudes between the mantle-folds, and thus converting it into a pearl, is strikingly illustrated by two specimens in which, in the one case, an entire fish, and in the other a small crab, have been so enclosed (see side table-case E).
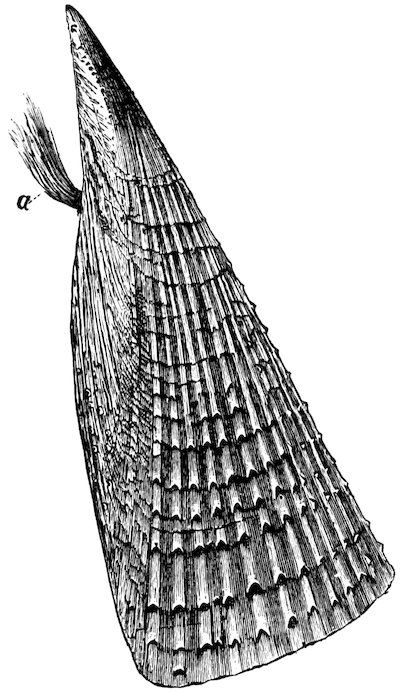
Fig. 28.
British “Fan-Mussel” (Pinna pectinata): a, the byssus. Case 150.
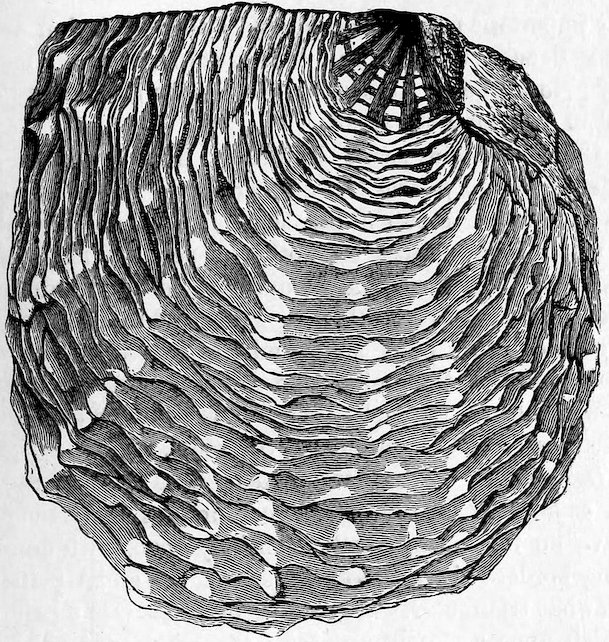
Fig. 29.
Pearl-Oyster (Meleagrina margaritifera). Case 146.
The most ancient and, even at the present day, one of the most important of the pearl-fisheries is that carried on on the western shores of Ceylon. “The Banks,” or spots on which the oysters grow, are at an average depth of 30 to 60 feet, and extend several miles along the coast. The oysters, which should be six or seven years old when collected, are gathered in baskets by native divers and hauled up by ropes into hundreds of small boats. The shells are then brought to land and placed upon the ground to die and putrefy, and then minutely examined for the pearls, which are either found loose in the shells or imbedded in the fleshy parts of the oysters. As many as two million oysters have been brought ashore on one day; but the number obtained varies very much according to the state of the banks. A small proportion of the oysters contain pearls; in some only very small ones (seed or dust-pearls as they are called) are found, and very few contain pearls larger than a pea, which are so highly valued. In his account of the 36pearl-fishery of Ceylon the Rev. James Cordiner says that he saw the operation of sorting the pearls performed; the produce of 17,000 oysters weighed only ¾ lb. and was contained in a vessel smaller than a common soup-plate. Out of that quantity there were not found two fine perfect pearls; all of the largest were slightly deformed, rugged and uneven, but of the smaller sizes many were round and perfect. The chief qualities which regulate the value of pearls are size, roundness, and brilliancy of lustre. Of the smallest kind several may be bought for a shilling, whilst many thousand pounds have been given for a single fine pearl of surpassing beauty.
Other important pearl-fisheries besides that of Ceylon are carried on in the Persian Gulf, on the west coast of Central America, and especially North-west Australia, where diving-dresses are now employed in collecting the shells.
The Chinese obtain pearls artificially from a species of freshwater Mussel (Dipsas plicata). In order to do this they keep them in tanks and insert between the shell and the animal either small shot or small round pieces of mother-of-pearl, which soon receive regular coatings of nacre and assume the look of ordinary pearls. They also insert small metal images of Buddha, which also soon become covered with pearl and firmly cemented to the shell, the production being to the uninitiated a supernatural testimony to the truth of Buddhism. (A shell treated in this way is exhibited in one of the small cases (E) at the side of the room.)
The Ostreidæ, or Oysters, undoubtedly take the first rank among molluscs as regards usefulness to mankind as an article of food. They have no foot; the mantle is entirely open, with double edges, each being bordered by a short fringe, and the labial palps are large and somewhat triangular. There are on each side a pair of simple gills, which appear closely striated; the single adductor muscle is large and nearly central (see Fig. 30). The Oyster is, except in the very young state, entirely incapable of locomotion, and always attached by the deeper valve to other shells, rocks, or other substances. The common British species is not full-grown until it is about five or seven years old. A series of different ages, from the “spat” to the adult form, is exhibited in Case 152. During the months of May, June, and July the eggs are discharged into the gills, where they remain until hatched; and it is during this period that oysters are “out of season.” In the American Oyster (O. virginica), on the contrary, the eggs are said to be 37hatched outside the parent shell. Oysters of different kinds are found on nearly every shore. The gigantic O. gigas is said to grow to the length of three feet in the Bay of Taichou, Japan, where it is commonly eaten. About two hundred fossil species have already been described.
The Spondylidæ, or Thorny Oysters, closely resemble the Scallops, but the shells are more spiny, heavier, united by interlocking teeth, and one of the valves is attached to rocks, corals, etc. Many of the species are very brightly coloured; and from the fact that small quantities of water are sometimes enclosed in cavities in the inner layer of the shell, they have been called “Water-Clams” or “Water Spondyli.”
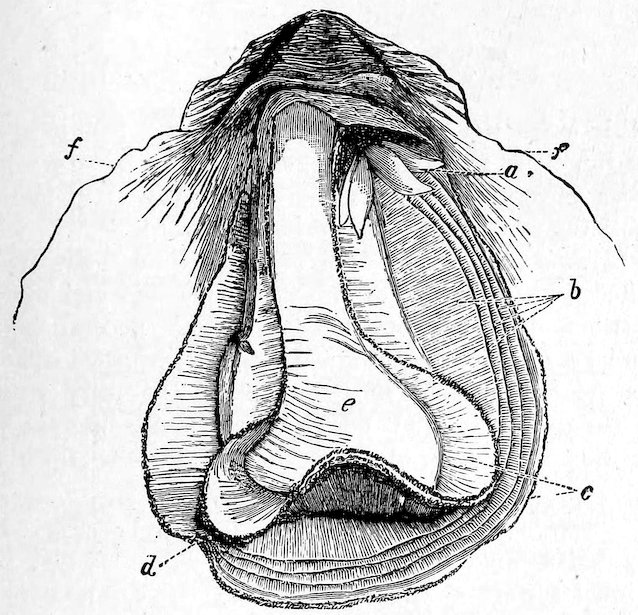
Fig. 30.
Common Edible Oyster (Ostrea edulis).
a, labial palpi; b, gills; c, mantle; d, junction of the two folds of the mantle; e, large adductor muscle; f, the shell.
The Limas (Limidæ) are very like the Pectens, but the inner edge of their mantle is fringed with very long thread-like filaments. The shells are always white, generally more or less oblique, and radiately ridged. They appear to be found in most seas, and either swim about freely like the young Scallops by flapping their valves 38or attach themselves by a byssus, sometimes forming a sort of nest, consisting of pieces of coral and shell or small stones, in which they are completely concealed.
The Scallops or Fan-shells (Pectinidæ) are well known for their beautiful colours, sculpture, and excellent flavour. The animal has a distinct foot, which is not, however, used as a locomotive organ, but employed in spinning a byssus of attachment when required. The young Pectens dart through the water by opening and suddenly closing their valves. The species are very numerous, world-wide in their distribution, and may be found at depths from a few to three thousand fathoms.
In this order the gills have interfilamentar and interlamellar vascular junctions, and there are generally two adductor muscles. The order is very extensive, comprising nearly sixty families, of which only the more important can be referred to.
The Carditidæ and Astartidæ have strong solid shells, frequently ornamented with radiating or concentric ribbing, and usually are coated with a dark epidermis. They have the general appearance of certain Veneridæ; but the animal has no prolonged siphons, but merely a fringed opening in the mantle. One very remarkable species, Thecalia concamerata (Case 161), has an internal cup-like process within the valves, which serves as a nursing-pouch for the young.
The Lucinidæ are almost invariably white shells, and may generally be recognized by the very long muscular scar in front on the inner surface of the valves. They occur in all parts of the world; and the fossil forms, which are still more numerous than those now living, have existed at every epoch from the Silurian.
Of the freshwater Mussels or Unionidæ more than 1200 species have been already discovered; they are found in most parts of the world, the greatest number having been described from North America. In Unio the edges of the mantle are not united along the bottom and not prolonged into siphonal tubes; at the posterior end there are two openings, of which the upper or excretal orifice is simple, and the lower or branchial fringed at the edge. Two “cephalic eyes” have recently been noticed by Mr. P. Pelseneer. The foot is very large and adapted for crawling and burrowing. The sexes are distinct; and the shells of the females are somewhat 39more tumid than those of the males. Margaritana margaritifera, (Case 168) which is found in this country and in Europe, sometimes produces handsome pearls, but not equal to those obtained from the pearl-oyster of tropical seas. The hinge in this family is extremely variable, being in some instances delicate and toothless (Anodonta, Fig. 31, B), whilst in others it is enormously thickened and furnished with strong interlocking teeth (see Fig. 31, A).
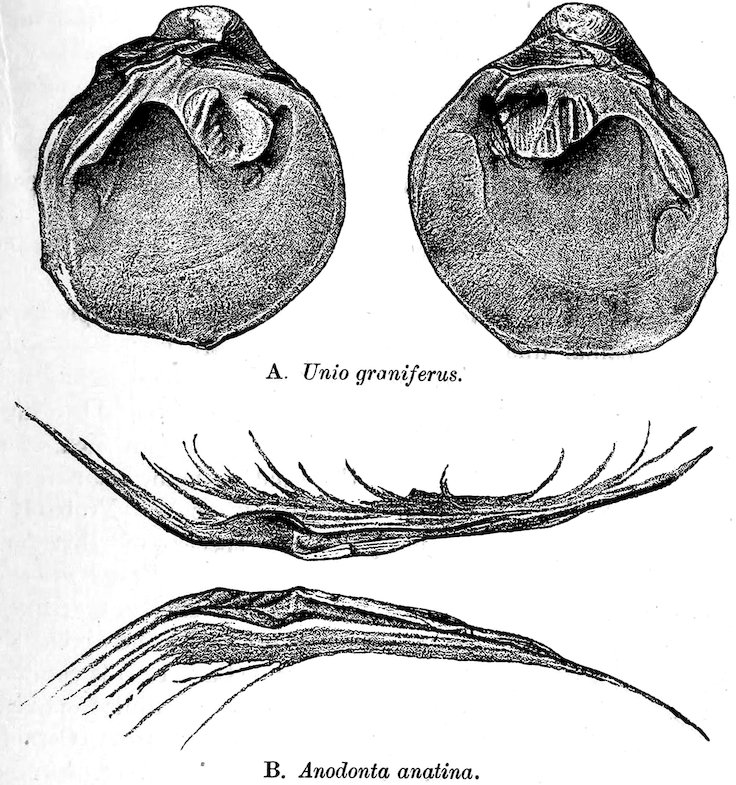
Fig. 31.
The family Ætheriidæ, or freshwater Oysters, consists of but three genera: Ætheria contains African, and Mülleria and Barttettia Indian and South-American forms. When young the shells of Ætheria (which are common in the Nile) are free and not unlike an Anodonta, but when adult they become attached and irregular and look like an olive-green Oyster; they are, however, provided with two muscular impressions instead of one, as in ordinary marine 40Oysters. Still more remarkable is Mülleria lobata of Colombia, which, when young, freely moves about and has two adductor muscles, but in time becomes attached and stationary, and then possesses but a single adductor.
The Tellinas have usually thin shells, and their two siphons are longer and more completely separated from each other than in the Veneridæ. The pallial line is widely and deeply sinuated, and the ligament generally external. In the genus Semele of the family Scrobiculariidæ, it is placed within the hinge-margin. They live in great numbers beneath the sand in shallow water, and are occasionally used as food.
The Mactridæ have an internal ligament to the hinge, the siphons are joined together and fringed at the ends, and the pallial line is more or less sinuated. Spisula solidissima (Case 184), the largest species found on the coast of the United States, is a common article of diet.
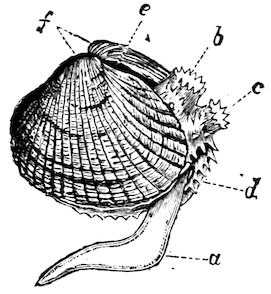
Fig. 32.
Common British Cockle
(Cardium edule).
a, foot; b, exhalant siphon; c, branchial or inhalant siphon; d, edge of mantle; e, ligament; f, umbones or beaks of the shell.
The next family, Veneridæ, have long respiratory siphons and a sinuated pallial line. Many of this tribe are very beautiful in form and colouring, and most of them have very hard strong shells. The valves are united above by an external ligament, and the hinge-plate is toothed. Nearly all of them live buried an inch or two beneath the sand or mud, but a few are found burrowing in rocks. Probably the majority of the species of this family might be used as food. Venus verrucosa, of our own southern shores, is frequently eaten both in this country and abroad; and Venus mercenaria (Case 189) is commonly sold in the markets of Philadelphia and New York. Cytherea lusoria (Case 185) also forms a favourite article of diet among the poorer classes in Japan, and several kinds are eaten by the natives of New Zealand and other countries.
Some of the “Cockles” (Cardiidæ) from warm latitudes are highly coloured and adorned with most beautiful sculpture. Probably the majority are eatable, as the common cockle (Cardium edule) of the British coast. The foot of these molluscs is very large, bent, and used for leaping. The siphons are short and fringed at the margins.
41The Tridacnidæ, or true Clams, differ from other Bivalves with united mantle-margins in having but a central adductor muscle. In the typical species the animal is attached to the rocks by a “byssus,” a strong fibrous structure which passes through an aperture at the upper part of the shell. A species found in the Red Sea, T. elongata, is eaten by the natives, and the shell employed for the manufacture of lime. Tridacna gigas, the largest known bivalved mollusc, sometimes weighs over 500 lb., that exhibited in the upright cases at the entrance of the Gallery being 310 lb. in weight. A large pair bordered with gilt copper are used as bénitiers or holy-water vessels in the church of St. Sulpice in Paris. Tridacnæ are found associated in large numbers in lagoons, among coral-reefs in the Eastern and Pacific Seas. The animals are described as presenting a beautiful iridescent glare of blue, violet, and yellow variegated with fantastic markings.
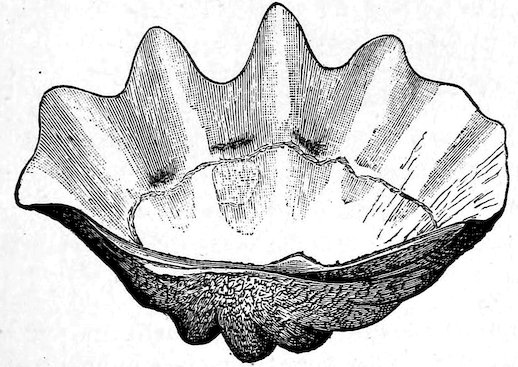
Fig. 33.
Left valve of the Giant Clam (Tridacna gigas).
Length, 36 inches. Weight, 154 lb.; weight of the two valves, 310 lb.
The genus Chama consists of tropical species, which are found fixed to corals, rocks, etc. Nevertheless, they have a small bent foot, but what purpose it serves is difficult to conceive.
In this place attention should be called to the Hippuritidæ and Radiolitidæ, very remarkable extinct families of bivalved molluscs which occur abundantly in the cretaceous strata of southern and eastern Europe, Egypt, etc. They are remarkable for the solidity of the shells, the relatively small space occupied by the animal, and the complicated character of the hinge and processes bearing the adductor muscles. They are usually classed near to the Chamidæ, but their true position as regards living Mollusca is very problematical. 42A fine series of these shells is exhibited in Gallery VIII., wall-case 5, in the Geological Department.
The Myidæ, popularly known as “Gapers,” on account of their valves being open at one or both ends, have the mantle united all round, except where the small foot is protruded. The siphons are very long, united almost to the ends, and covered with a coarse wrinkled outer skin. They bury themselves in mud and sand at low-water mark or in shallow water. The species are few in number, and chiefly from the shores of northern countries. Mya arenaria of our own coasts is largely eaten in some parts of Europe and North America.
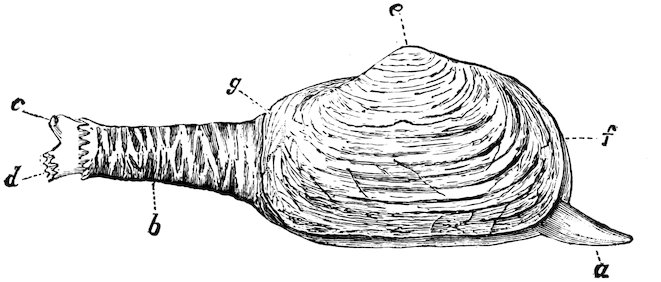
Fig. 34.
British Gaper (Mya truncata).
a, foot; b, siphon-sheath; c, exhalant siphon; d, inhalant siphon; e, umbones or beaks; f, anterior, g, posterior end of shell.
The Corbulæ (Case 198) have one valve larger than the other and are like little Myæ, but the valves are almost closed and their siphons are very short.
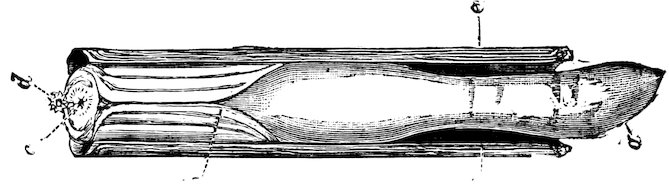
Fig. 35.
British Razor-shell (Solen siliqua).
a, foot; b, mantle; c, inhalant siphon; d, exhalant siphon; e, shell.
Many of the Solenidæ, or Razor-shells, possess very elongated shells, and are remarkable for the great development of the foot, which can be pointed or contracted as may be required for boring into sand. By means of this powerful foot the animals, when 43disturbed, bore with such rapidity and to such a depth that their capture is a matter of great difficulty; and even when seized they hold on so tightly that at times they suffer their foot to be torn off rather than be captured. They not only burrow in sand, but also have the power of darting through the water, like the Scallops. Solens were considered a dainty dish by the ancient Greeks, and numbers are still eaten by the poorer coast-population of this country and abroad.
The Pholadidæ, or Piddocks, are very remarkable shells, of an unusually complicated structure, some having the power of boring into rocks, wood, mud, sand, etc. Their shells are white, adorned with prickly sculpture, and, although thin, are strong. The foot is believed to be the principal excavating instrument, but the shell no doubt is used as a file to enlarge the hole as the creature grows. These animals are brightly phosphorescent; and certain species are eaten at many places on the shores of the Mediterranean. They appear to be indifferent as regards the material they bore into; for the common Pholas dactylus (Fig. 36) of our own shores has been found in slate-rocks, mica-schist, coal-shale, new red sandstone, chalk, marl, peat, and submarine wood. The siphons are long in the Piddocks, united except near the end, and enclosed in tough skin. The species are world-wide in their distribution, and several are found fossil in some of the Tertiary formations.
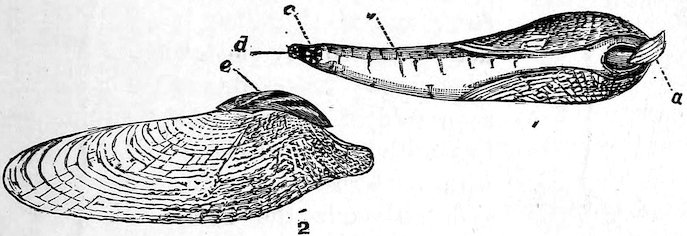
Fig. 36.
Piddock, or Borer (Pholas dactylus). (From the British coast.)
1. Animal in the shell: a, foot; b, siphons; c, inhalant orifice; d, exhalant orifice.
2. Shell: e, accessory valves or plates.
The Teredinidæ, or Ship-worms, are also borers, like the Pholads, but do not perforate rocks. They are principally wood-borers; the large Kuphus arenaria, which is an exception, living buried in the sand. The ship-worm has a long worm-like body, from 6 to 12 44inches in length, which is more or less enclosed in a thin shelly tube or sheath. The true bivalved shell is at the thicker end, and protects the mouth, labial palps, the liver, and other internal organs. At the opposite, or more slender, end of the animal, the mantle is produced into two small tubes, one of which conveys the water to the gills, whilst through the other the water is expelled, charged with the woody pulp excavated by the foot. At the end there is a pair of pallets, or paddles as they are sometimes termed, which are probably used as a means of defence, in closing the shelly tube after the contraction of the siphons.
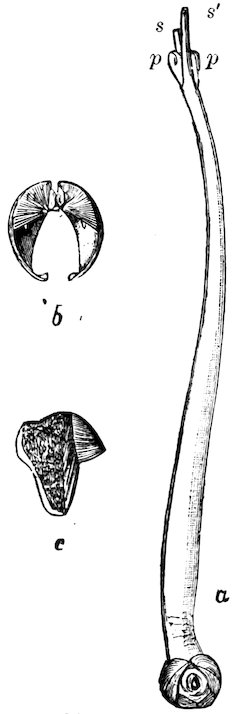
Fig. 37.
Ship-worm
(Teredo norvegica).
Case 202.
a, animal, removed from its shelly tube: p, p, pallets; s, exhalant siphon; s′, inhalant siphon.
b, c, different aspects of the shell.
These animals are most destructive to ships, piers, etc.; and wood, which is not protected by metal, when once attacked, is soon riddled through and through. They work either with or across the grain, and although the holes may be all but touching, they seldom appear to run into one another.

Fig. 38.
Watering-pot Shell
(Brechites vaginifer).
Case 204.
a, bivalve shell of the very young animal.
The “Watering-pot shell” (Brechites), of the family Clavagellidæ, is a very remarkable structure, and unlike the shell of an ordinary bivalved mollusc. On looking carefully, however, near the perforated end (the rose), two small valves will be seen imbedded in the surface. They are found with the rose downwards buried in mud or sand at low water on the shores of the Indian and Pacific Oceans.
The members of this order differ from other Lamellibranchs in having the gill-plates represented by a muscular septum. They are provided with two respiratory siphons and two adductor muscles, and the edges of the mantle-lobes are connected at three points. The families Poromyidæ, and Cuspidariidæ, constitute this order. The species are all small, without colour-markings, are world-wide in their distribution and occur at all depths.

Fig. 39.
A, the upper, B, the lower beak of Architeuthis monachus; one-third natural size.
This Class includes the Octopus or Polypus, Cuttlefish, Squid, Spirula, the Paper and Pearly Nautilus. The body of the animal consists of a muscular sac, in the cavity of which the viscera are placed. In front of the body projects the head, which, in species belonging to the two-gilled section of the Class, is surrounded by eight or ten fleshy arms. A wide aperture below the head admits the water to the gills or branchiæ, which are situated in the interior of the sac, whilst a short tube, the so-called funnel or siphuncle, projects from the opening of the mantle—the water and various excretions being expelled through this tube, especially also an ink-like fluid, which is discharged by all Cephalopods (except Nautilus) when disturbed, in order to darken the water and thus escape their enemies. The centre of the head, between the base of the arms, is occupied by the mouth, which is armed with two horny or calcareous jaws, similar to the beak of a parrot (Fig. 39). The two large eyes are placed on the sides of the head. The arms or feet are more or less elongate, capable of movement in any direction, and, except in Nautilus, furnished on one side with numerous suckers, by means of which the 46animal attaches itself most securely to anything it may seize; they are employed in capturing food and in walking. Cephalopods walk in any direction head downwards, but can swim backwards only, being propelled in that direction by the water which they discharge with force through the funnel out of their branchial cavity. They are divided, according to the number of their gills (which is either two or four), into Dibranchia and Tetrabranchia. Of the latter but one representative now exists, viz., the Pearly Nautilus, all other living Cephalopods being provided with but two gills, placed one on each side of the body within the mantle, as may be seen in the wax model of Sepia officinalis (Case 207). The two-gilled section comprises forms with eight arms, as Argonauta and Octopus, and others with ten arms, viz., the Cuttlefishes (Sepia) (Fig. 43), the Squids (Loligo, Ommatostrephes, Sepiola, Chiroteuthis, etc.), and Spirula. The “shell” of the Paper-Nautilus, or Argonauta, is too well known to require any description. Unlike the shells of other Mollusca, it is not attached to the animal by a special muscle, but is held on to the body by two of the arms, which are dilated and specially adapted for this purpose. Only the female Argonaut is provided with a shell, the male being shellless and a much smaller creature. The Argonaut-shell is therefore not a true shell, but simply a receptacle for the ova, serving at the same time for the protection of the parent.

Fig 40.
The Common Octopus (Polypus vulgaris), resting.
Fig. 41.
Sepiola scandica
(Natural size). British.
47Chiroteuthis Veranyi is remarkable on account of the great length of the tentacular arms. These are non-retractile and are employed to seize their prey when at a distance.
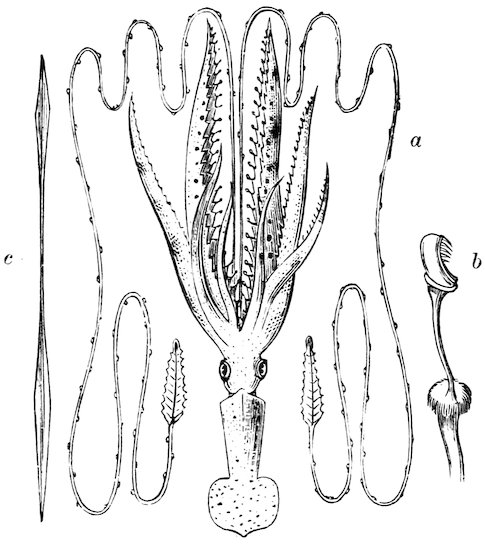
Fig. 42.
Chiroteuthis Veranyi (much reduced).
a, general view of animal; b, magnified view of pedunculated sucker of the terminal club of the tentacular arms; c, internal shell or gladius.
The species of Octopus are found on the shores of almost all temperate and tropical seas; they do not attain to a large size, and are without the internal shell or “bone” which is found in the mantle of many Cephalopods. That of the Cuttlefish or Sepia (Fig. 43 a) is found in abundance on our coasts; it is composed of numberless layers of a friable calcareous substance. That of the Squid tribe is of quite another character, consisting of an elongate thin horny plate, and strengthened by one or more thickened ribs, in some species somewhat resembling a quill-pen. Some species of this pen-bearing class related to the Common Squid attain an immense size. 48One was captured off the Irish coast in June, 1875 (probably Architeuthis harveyi), with the shorter arms 8 feet in length and 15 inches in circumference at the base, the two tentacular arms having a total length of 30 feet. The powerful beak measured about 4 inches across. Thus from the tip of the tail to the end of the tentacular arms this wonderful monster must have measured something like 40 feet in length. Other very large specimens of Architeuthis have been captured on the coasts of Newfoundland and Labrador. Two specimens stranded on the south coast of Newfoundland, in the winter of 1870–1871, measured respectively 40 and 47 feet. Another, cast ashore at Bonavista Bay in December, 1873, had a very stout body 14 feet long, arms 10 feet, and tentacles 24 feet in length. These are only a few of the many instances of the capture of gigantic Cephalopods, which occur not only in the North-Atlantic Ocean, but also in tropical seas. Their appearance in mid-ocean may, in some instances, have given rise to the tales of “Sea-serpents.” Specimens much smaller than those mentioned above have attacked men, and pearl-fishers are in constant fear of them. One of the arms of a large Squid (Architeuthis harveyi?), which is supposed to have been found off the coast of South America, is exhibited in the black upright Case A at the side of the room.
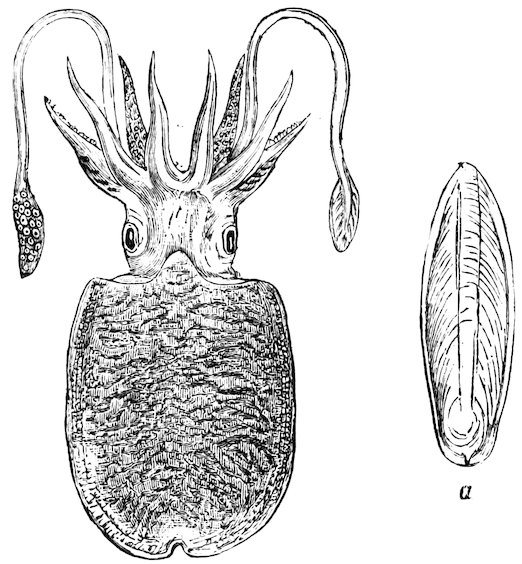
Fig. 43.
The Common Cuttlefish (Sepia officinalis), and its shell or bone (a).
49The shells of Spirula (Fig. 44) have been long known, and are scattered in thousands on the shores of New Zealand and other islands in the Pacific Ocean, and they are also found in the Indian and Atlantic Oceans, occasionally drifting on the coast of Devon and Cornwall. Notwithstanding the abundance of the shells, very few specimens of the perfect animal have been captured. The loosely-coiled shell resembles a ram’s horn, and is divided into a number of segments by fine concave partitions, like the shell of Nautilus, each one pierced by a slender tube or siphon. It is placed at the hinder end of the body, and is covered with so thin a skin, that a small portion of it appears to be exposed both in front and behind. Absolutely nothing is known of the habits of this very interesting creature, although probably they are somewhat similar to those of other Cephalopods.
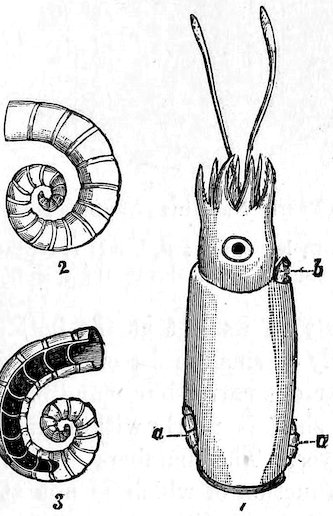
Fig. 44.
The Spirula (Spirula peronii). (From the Indian and Pacific Oceans.)
1. Animal: a, portions of the shell exposed in front and behind; b, the funnel or siphuncle. 2. Side view of shell. 3. Shell in section, to show partitions or septa.
The Nautilus (Fig. 45), of which several shells (Case 208) and a perfect animal in spirit (black upright case A) are exhibited, is an inhabitant of the Indo-Pacific Ocean, and differs from all other living Cephalopods in being provided with four instead of two gills, and, instead of eight or ten arms with suckers and hooks, has a 50number of small retractile feelers. The Nautilus occasionally swims, like other members of its class, at the surface of the sea, but mostly crawls about leisurely on its feet at the bottom in search of food, which consists chiefly of small crabs or Mollusca, which it crushes with its strong calcareous parrot-like mandibles.
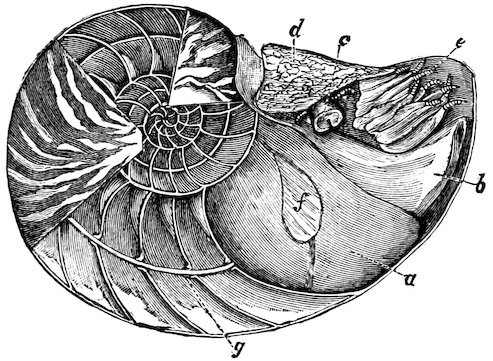
Fig. 45.
The Pearly Nautilus (Nautilus pompilius).
a, body; b, siphuncle; c, eye; d, hood; e, tentacles; f, muscle of attachment to the shell; g, siphon.
The chambered shell is pearly within, and covered with an external calcareous layer. The chambers are connected by a slender tube or siphon, the function of which is not at present thoroughly understood. The septa, or partitions across the shell, indicate periods of growth. When the Nautilus outgrows the capacity of the outer chamber, in which it resides, it constructs a new one of larger size, separating the additional chamber from the preceding one by a transverse partition.
A series of Cephalopods preserved in spirit is exhibited in the black upright case at the side of the room.
This Index has been compiled to assist the numerous visitors, who wish to examine and determine specimens of shells, in finding, without trouble or loss of time, the Cases in which the genera are placed. Subgeneric terms are omitted, as they do not fall within the scope of this “Guide.”
From a casual glance at the contents of these cases, it might be supposed that many of the specimens exhibited therein were seaweeds; but a closer inspection, especially with a lens, will reveal structure of a kind not to be found in any plant.
Let us select for examination Flustra foliacea, the Broad-leaved Hornwrack or Sea-Mat (Fig. 1), (Case A 1), commonly to be found among heaps of seaweed cast up on sandy shores round our coasts.
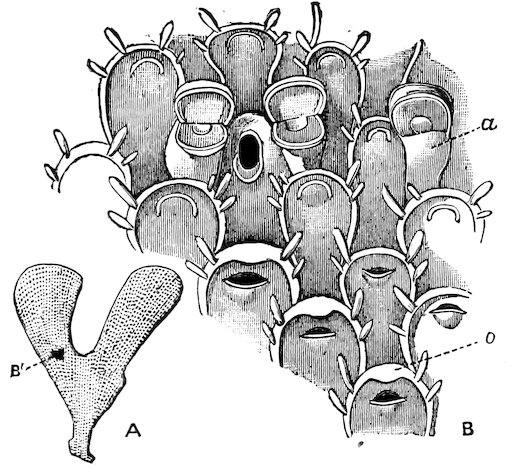
Fig. 1.
Flustra foliacea. A, natural size; B′, portion magnified in B; B, magnified 30 diameters.
a, avicularium; o, ovicell.
[‘The Cambridge Natural History.’]
The brown horny fronds, which vary in width, branch upwards from a narrow flat stem attached at its base to stones and shells. Both surfaces of the fronds show a fine network pattern formed by 55the edges of little oblong boxes or cells termed zoœcia,[10] arranged in longitudinal parallel rows and forming a double layer back to back. The cells are broad and rounded above, narrow and truncate below, and each is roofed in by a transparent membrane with a semicircular lid or operculum situated near the upper end; four short stout spines spring from the margin in this neighbourhood. When the surface of a living frond is examined in sea-water, here and there a bundle of tentacles may be observed pushing up a lid, slowly emerging and expanding into a bell-shaped coronet; on the least alarm the tentacles are rapidly withdrawn into the cell and the lid shut. The flexible protrusible region of the cell is termed the tentacle sheath. The relation of the cell to the tentacle sheath (Figs. 2, 3) may be roughly compared to a glove finger, stiff below, but flexible at the end, and surmounted by a crown of bristles; on pulling down the glove-finger tip, the tentacles will also be drawn in, and will lie in a sheath formed by the invaginated portion of glove finger. The lid which closes over the tentacle sheath is only found in the Sub-order Chilostomata to which Flustra belongs. The area of the tentacle sheath whence the tentacles arise is termed the lophophore.[11]
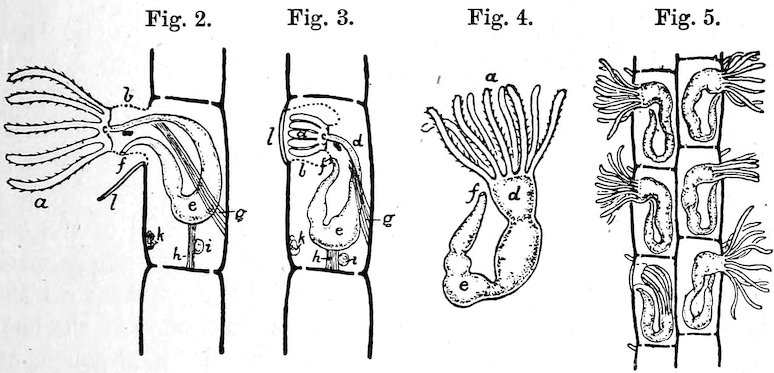
Figs. 2, 3, diagrams representing polypide in cell. Fig. 2, tentacle-sheath protruded. Fig. 3, ditto, retracted; a, tentacles; b, tentacle-sheath; c, mouth; d, gullet; e, stomach; f, vent; g, retractor muscle; h, funiculus; l, ovary; k, testis; l, lid or operculum; nerve ganglion is between mouth and vent. Fig. 4, polypide extracted from cell; d, pharynx; e, stomach; f, anus (after Van Beneden). Fig. 5, section (partly diagrammatic) of frond of Flustra, showing cells back to back.
56The mouth is situated in the centre of the lophophore, surrounded by the circle of tentacles; and the latter, by the action of their cilia, set up currents which convey food to the mouth.
The mouth leads into a pharynx and gullet, the latter opening into a stomach, whence the intestine ascends to terminate in the vent opening below and outside the circle of tentacles; the intestines, in fact, form a U-shaped tube (Figs. 4, 5) suspended in the body-cavity in the interior of the cell. A cord, the funiculus, passes from the stomach to the base of the body-cavity. A small nerve ganglion is situated within the upper part of the loop of intestine.
The tentacles, intestines, and other organs constitute the “polypide,” the cell being simply the protective house formed by the latter.
The body-cavity, which contains fluid, is in direct communication with the interior of the tentacles, which are hollow, and which act as respiratory organs by bringing the fluids of the body-cavity in proximity to the water. In Flustra the body-cavities of the cells are shut off from each other, but pores and sieves in the partition walls allow of the junction of the inner linings of these cavities. The male and female reproductive elements are formed in the body-cavity. The egg develops in a helmet-shaped brood-pouch, the ovicell, situated at the summit of the cell and almost immersed in the cell above. The ciliated embryo swims about for a few hours and settles down to form the first polypide and cell; from the latter there arise buds which remain attached, and produce other buds, till a colony like that of Flustra results.
Among the ordinary cells are certain smaller cells (Fig. 1, a) slightly raised above the general level, different in shape from the ordinary kind and with thicker lids. These peculiar cells are termed avicularia, and chiefly contain muscles for opening and shutting the lid. They arise by modification of the ordinary cells, whereby all the organs of the polypide have become atrophied except the muscles. The Polyzoa[12] were so named by Vaughan Thompson, who, in 1820, discovered that certain plant-like animals, which had previously been classed with the zoophytes, possessed a much higher organisation, in that the intestine was separate from the body-cavity and not continuous with it as in Sea-Firs, Sea-Anemones, and Corals. In 1834, Ehrenberg named the group Bryozoa[13] or Moss Animals.
57With the exception of one genus (Loxosoma), all Polyzoa form colonies, which arise by the continual budding of the cells, the buds remaining attached to the parent cells. The colonies vary endlessly in form and habit, occurring as crusts on rocks, etc., masses, broad fronds, branching tree-like growths, bushy tufts, etc.
The texture and consistency may be gelatinous, cartilaginous, horny and flexible, or stony.
The great majority of species are marine, but a considerable number inhabit fresh water. The Polyzoa are classified as follows:—
The Chilostomata, which contain many more species than all the other groups put together, are divided into three sections:—A. Cellularina, in which the cells are more or less boat-shaped or cornucopia-shaped, and joined together to form flexible branching colonies; B. Flustrina, in which the cells are typically shaped like oblong boxes with membranous front walls; and C. Escharina, in which the whole front wall is calcified.
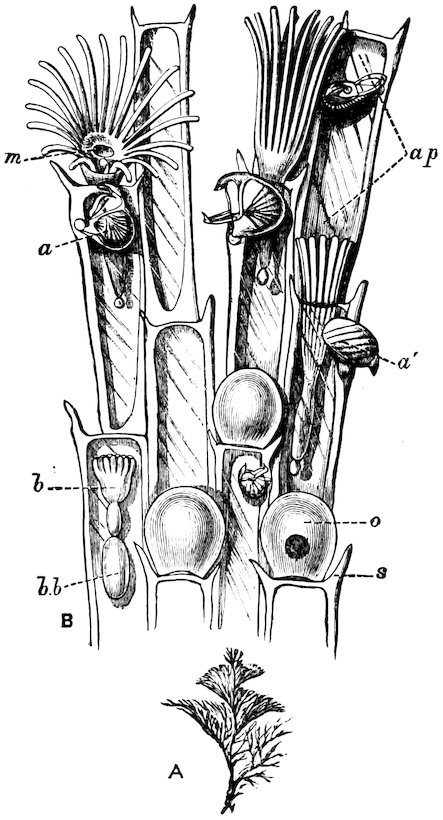
Fig. 6.
A, Bugula turbinata, natural size, B, portion × 50.
a, avicularia; m, mouth; o, ovicell.
[‘The Cambridge Natural History.’]
Section A. Cellularina.—Bugula turbinata, or the Bird’s-head Coralline (Fig. 6) grows attached to rocks near low water mark in the form of spiral tufts about two inches in height, composed of narrow flat branches in which the cells are arranged from two to six abreast and all facing upwards. Each cell is boat-shaped and with nearly the whole front surface membranous; the globular bodies at the head of certain cells are the ovicells. Attached to the outer edge of each cell is a remarkable object resembling a bird’s head, and hence termed avicularium, seated on a short stalk. The head and beak contain powerful muscles for opening and shutting a horny lid 59or mandible hinged on below. In life, the avicularium sways to and fro on its stalk, with the lower “jaw” continually snapping up and down in the most ludicrous fashion. The beak is capable of seizing and holding quite large objects.
The function of these curious appendages is partly to warn off trespassers and partly to capture and retain small animals till decomposition has set in; in the latter case, the currents set up by the tentacles draw in the particles to the mouths of the polypides. The avicularia have arisen by modification of the ordinary cells, in which the muscles have developed at the expense of the degenerated polypides, the cells have become much smaller, of different shape, and separated out from the rest; the mandible represents the lid or operculum of the ordinary cell. The avicularia vary greatly in size and shape in the different genera; in Flustra, for instance, these organs closely resemble the ordinary cells.

Fig. 7.
Bugula bicornis. Cells magnified. (After Busk.)
In Bugula bicornis(*) (Fig. 7), from 1950 fathoms in the Southern Indian Ocean, each cell is provided with two avicularia with remarkably long stalks. The graceful vase-shaped Kinetoskias cyathus(*) 60(Fig. 8), one of the treasures of the “Challenger” Expedition, was dredged from 1525 fathoms off Cape St. Vincent. The stem, which tapers gradually upwards, rises from a tuft of root fibres. The cup is formed of slender branches supported at the base by a delicate membrane. The branches are composed of biserial rows of cells (Fig. 9) opening towards the interior of the cup. The avicularia are pear-shaped and pedunclate. Probably, in life, the cup is capable of being opened out to a considerable extent. Specimens of this species were also obtained from 2160 fathoms in the South Atlantic.
Fig. 9.
Kinetoskias cyathus. A branch magnified.
a, an avicularium. (After Busk.)
Scrupocellaria reptans, or the Creeping Coralline (Fig. 10 A, B) forms branching colonies, creeping over rocks and seaweeds, and attached by horny fibres often provided with curved hooks. The branches are composed of cells arranged in a double row. Each cell has the membranous area of its front surface protected by a branched flattened spine or operculum, and is produced and narrowed below; at the upper outer margin is a minute triangular avicularium. At the base of the back surface is a small sack-shaped cell with a cleft at the upper end, in which a horny bristle is articulated. The little cell is termed a vibracular cell, and the bristle a vibraculum.[21] This organ has arisen by a further modification of an avicularium, whereby the horny lid of the latter has become a long bristle. The bristles by their motion keep off intruders, and possibly act as scavengers by sweeping the surface of the cells.

Fig. 8.
Kinetoskias cyathus. (From Voy. Challenger, Atlantic: Wyv. Thomson.)
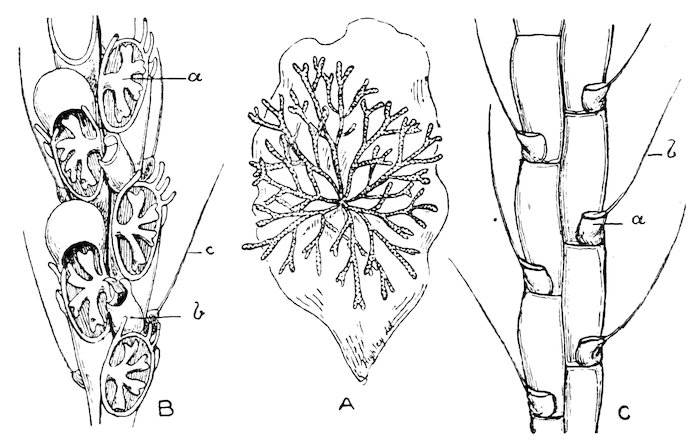
Fig. 10.
Scrupocellaria reptans. A. Creeping over seaweed, natural size; B. Front surface, magnified.
a, branched spine covering front of membranous area; b, avicularium; c, vibraculum.
C. Back surface; a, vibracular cell; b, vibraculum.
In Caberia ellisii the vibracular cells are very large. The vibracula, which are long and serrated, have been observed to move in unison like a double row of oars.
Section B. Flustrina.—In this group the colonies form leafy lamellæ, crusts, etc., in which the individual cells are typically in the form of oblong boxes with their front walls wholly or partly membranous. Flustra foliacea has already been described. |Case A 1.| In Flustra carbasea the fronds are formed of only one layer of cells, and not of two layers back to back as in F. foliacea. The fine specimen of Flustra nobilis from S. Africa is so called from the large size of its long hexagonal cells which form a honeycomb pattern clearly visible to the naked eye.
In Flustra cribriformis(*) (Fig. 11), from Torres Straits, the fenestrated frond forms a beautiful spiral. Flustra florea, from S. Australia, grows in the form of branching tufts of narrow spiral fronds. Electra pilosa [dry and spirit specimens exhibited] (Fig. 12) 63forms a delicate silvery lace-work, encrusting shells and seaweeds (especially red algæ) on almost every shore. The long horny spine at the base of the membranous area of each cell gives the crust a pilose appearance. In Electra verticillata from West Africa, the cells form an elegant branched colony, the branches being composed of regular verticils of cells.
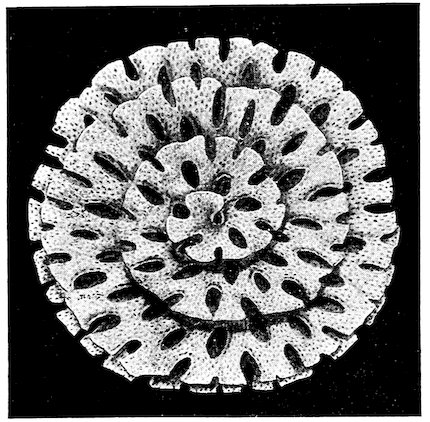
Fig. 11.
Flustra cribriformis.
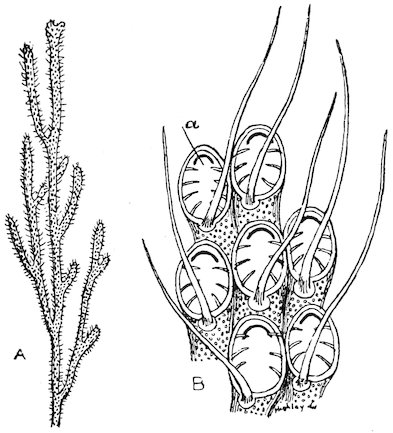
Fig. 12.
Electra pilosa. A, incrusting a seaweed, natural size; B, cells magnified; a, lid or operculum.
Membranipora membranacea occurs in the form of horny incrustations 64on bladder-wrack, which, owing to their flexibility, are able to adapt themselves to the swaying of the fronds of the Fucus.
The Selenariidae (Case B 2) form free colonies, usually orbicular in shape, convex above and concave below. In Lunulites capulus alternating rows of cells and vibracula radiate from the centre of the colony.
Section C. Escharina.—In this group, the front walls of the cells are wholly calcareous. Many species form patches or crusts on shells etc., and hence the name of the section; other species, again, form stony tree-like growths, or thick plates. Frequently one and the same species occurs in the form of crusts or of erect lamellæ, the identity being recognised by the characters of the individual cells.
Often a large number of species may be found on one shell. Two good examples of this are exhibited in Case A 2.
Lepralia pallasiana (Fig. 13) forms sub-circular vitreous patches on stones and shells; the cells are rather large, broadly oval, and with the front wall punctured with pores; the aperture is squarish and with a slight indentation on each side.
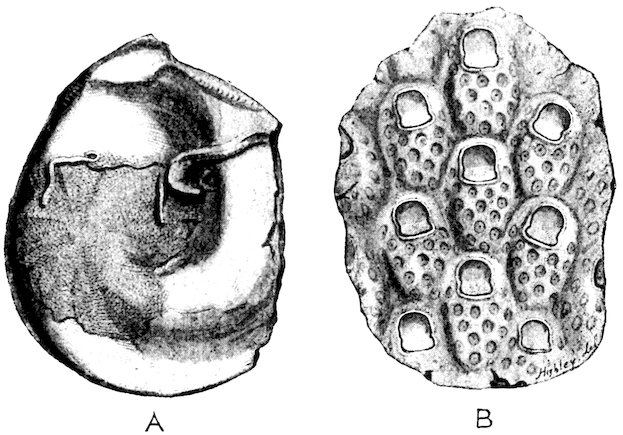
Fig. 13.
Lepralia pallasiana, incrusting a shell. A, natural size; B, cells magnified.
Lepralia foliacea forms a massive coral-like growth composed of thin contorted plates which fuse to form labyrinthine cavities, the plates being constructed of a double layer of cells back to back. A large specimen from the English Channel is exhibited in Case B, upright part. In Lepralia the orifice and lid of the cell have a straight lower margin, but one large group, Myriozoidæ, is characterised 65by having a notch in the lower margin of the orifice, (Fig. 14, Schizoporella unicornis).
In many of the Escharina, the front wall of the cell is produced into a stout process or mucro at the lower margin of the orifice (genus Mucronella), or, again, a collar or tube grows up round the primary orifice, thus giving rise to a secondary orifice (Smittia, Porella, etc., Case B 1).
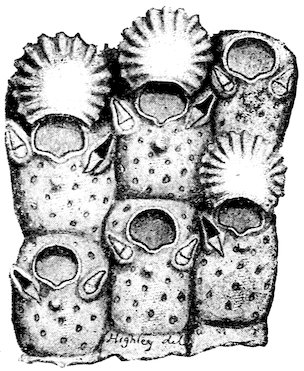
Fig. 14.
Schizoporella unicornis, magnified.

Fig. 15.
Retepora beaniana.
In the Celleporidæ, (Case B 1) the cells are typically pitcher-shaped and arranged vertically, and tend to be heaped up from the overcrowding.
Cellepora pumicosa forms thick pumice-like masses composed of 66succeeding layers of cells. The Reteporidæ (Case B 1) form delicate stony networks. The reticulate fronds may be expanded out, or may form tubular or contorted growths (Fig. 15, Retepora beaniana). The beautiful Retepora phœnicea from Torres Straits is of a rich purple colour.
The Adeonidæ form thick fenestrated plates which unite to form cavernous masses usually attached to rocks by a thick jointed stem. Several very fine examples from Port Phillip, Victoria, are exhibited in the upright part of Case B.
The Catenicellidæ are represented by a fine series of specimens from Australia. The colonies form dense clusters of finely beaded branches. The cells are arranged in single series, each cell being united to those above and below by a horny joint. The cells are usually urn-shaped with a triangular avicularium at each upper angle, and with the front surface variously sculptured with pores or bands (Fig. 16, Catenicella ventricosa).
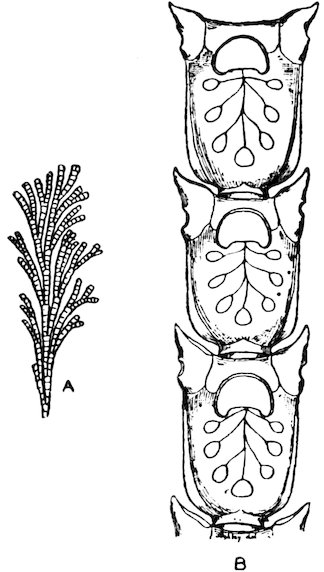
Fig. 16.
Catenicella ventricosa. A, natural size; B, magnified. (After Busk.)
The Ctenostomata are fleshy, horny, or membranous; never calcareous. When the tentacles of a polypide are retracted into 67the cell, they are protected above by a membranous comb-like frill.
The cells either bud off from each other or arise as buds on a stolon or stem.
Alcyonidium gelatinosum(*) (Fig. 17), so called from its resemblance to the zoophyte Alcyonium, forms fleshy translucent growths occurring in the form of nodulated branched masses, or of long finger-like growths. The species is common round our coasts where it grows attached to stones and shells near low-water mark.
Fig. 17.
Alcyonidium gelatinosum. A, a small piece, natural size; B, the same magnified.
Amathia forms bushy growths composed of slender horny branches. The cells, which are cylindrical or squarish, rise from the branches in biserial rows like Pan’s pipes. In Amathia lendigera(*) (Fig. 18) the groups of cells are well separated from each other, but in A. spiralis(*) and A. convoluta(*) the cells form a nearly or entirely continuous series winding in a spiral round the slender stems. In Bowerbankia imbricata(*) the cells are clustered on the stems. Vesicularia spinosa(*), 68or the Silk Coralline, forms delicate brown tufts resembling a filamentous alga; the cells arise separately in a single series from the hollow tubular stems and are contracted at their point of attachment.
Fig. 18.
Amathia lendigera. A, natural size; B, magnified.
Nearly all the Ctenostomata are marine, but a few species live in fresh water.
In the Cyclostomata, which are all calcareous, the usually tubular zoœcia have plain circular orifices without a lid or frill closing over the retracted tentacle-sheath. There are two sections in this group, viz., Articulata, in which the cells form branching colonies, the branches being connected by horny joints; and Inarticulata, 69in which the colonies may be encrusting, or erect and branching, but are without joints.
The first section includes the Crisiidæ.
Crisia denticulata (Fig. 19) forms delicate white tufts, in which the flat slender branches are composed of a double row of tubular cells. The horny joints between the branches are black. The Inarticulata occur as crusts or branching growths. In Tubulipora flabellaris (Fig. 20) the colonies form little fan-shaped crusts on seaweeds. Lichenopora hispida forms little white disks, in which rows of tubular cells radiate from the centre. In Idmonea, the colony is branched, the tubular cells being arranged in parallel rows on each side of the middle line of the branch.

Fig. 19.
Crisia denticulata. A, natural size; B, branches magnified.
The Cyclostomata are all marine.
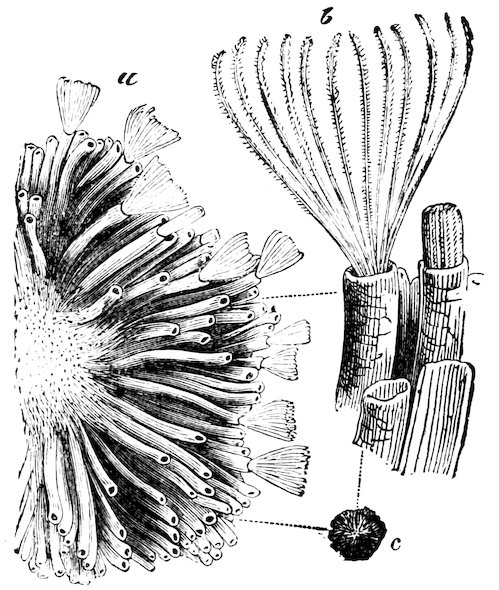
Fig. 20.
Tubulipora flabellaris.
a, half of an incrusting colony, × 8; b, a few cells, × 44; c, a colony, natural size.
All the forms in this group inhabit fresh water, where, in the form of creeping or erect branching growths or masses, they grow attached to freshwater plants, tree-trunks, old wood, etc.; two species are capable of slow movement from place to place. The lophophore and tentacular crown of the polypide are horseshoe-shaped. The Order owes its name to the presence of a lobe guarding the mouth.
In addition to the sexual, there is an asexual reproduction by means of peculiar internal buds termed statoblasts (Fig. 21). When the colony dies in the autumn, the liberated buds, securely protected in a horny capsule, retain their vitality till the spring; in due season the valves of the statoblast burst open, and the contents develope into a new colony. The statoblasts, which resemble small seeds, are usually provided with a ring of air cells, which act as a float, and in some species spines are present.
Fig. 21.
Statoblasts of Freshwater Polyzoa. A, Fredericella sultena × 38; B, Plumatella repens × 38; C, Lophopus crystallinus × 28; D, Cristatella mucedo × 28.
[‘The Cambridge Natural History.’]
Fig. 22.
A, Plumatella repens, partly free, partly incrusting stem of water-weed. B, Cells magnified. (After Allman.)
72Plumatella repens(*) (Fig. 22) forms brown branching colonies, wholly or partly adherent to the surface of leaves of water plants, old wood, etc. The individual cells are club-shaped, and about a quarter of an inch long, each cell being attached to the upper back part of the cell below; the statoblasts (Fig 21, B) are simple oval bodies with a zone of air cells.
Plumatella (Alcyonella) fungosa forms thick masses, composed of closely packed vertical tubes. A small specimen(*) surrounding a stick from Hampstead Ponds is exhibited.
Lophopus crystallinus(*) occurs in the form of translucent gelatinous blobs, often attached to the slender stems of duck-weed. The statoblasts (Fig. 21, C) are elliptical and pointed at each end. The polypides are comparatively large, and can be easily observed through the transparent surface. When its delicate plumes are fully expanded, Lophopus forms a beautiful object.
Cristatella mucedo(*) (Fig. 23) occurs in the form of greenish translucent oval or worm-like colonies with the polypides on the convex upper surface. The animal slowly creeps about on its flattened under surface.
Fig. 23.
Cristatella mucedo, creeping over a stem of water-weed; × 6. (After Allman.)
a, polypides with horseshoe-shaped crown of tentacles; b, statoblasts seen through the tissues; c, muscular sole by means of which the animal creeps; d, stem of water-weed.
Freshwater Polyzoa usually prefer dark places, but Cristatella creeps along on the stones and pebbles in clear water, and in the sunlight. The polypides form three or more concentric rows on the upper surface. The statoblasts (Fig. 21, D) are circular, provided 73with a zone of air cells, and with hooked spines, the total diameter being about ¹⁄₃₀ of an inch.
In this small group, both orifices of the alimentary canal open within the circle of tentacles, and there is no tentacular sheath.
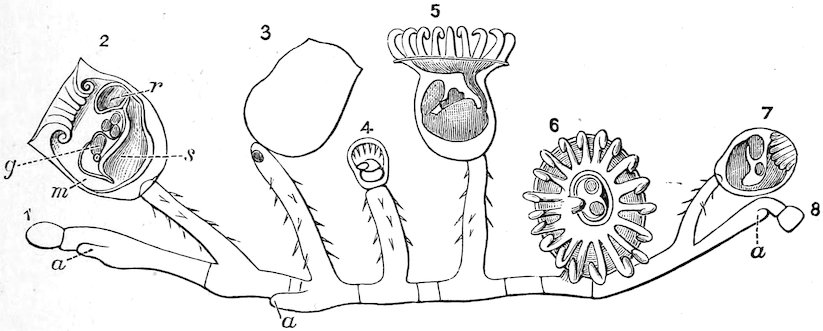
Fig. 24.
Pedicellina cernua. × 27.
[‘The Cambridge Natural History.’]
The polypides are borne on contractile stalks. In the Pedicellinidæ the stalks arise from a creeping stolon. In Pedicellina cernua(*) (Fig. 24) a stolon, creeping over seaweeds, etc., gives rise to stalked cups, the movements of which are vigorous: “the polypides, when excited, dash themselves vehemently from side to side. The heads are easily knocked off, but the decapitated stalks develop fresh ones. In Ascopodaria the stalks are swollen at the base; A. fruticosa(*), from Port Phillip, Victoria, forms beautiful tree-like colonies. The Loxosomidæ do not form colonies, owing to the buds becoming detached from the parent. The species of Loxosoma are always found associated with some other animal, such as a worm or Tunicate. The tentacles of the polypide are arranged obliquely to the long axis of the body, hence the name of the family (loxos, oblique). Loxosoma phascolosomatum(*) occurs, in the form of delicate tufts, on the caudal end of the Sipunculid worm Phascolosoma. The individuals resemble pins with little white heads, and are capable of vigorous movements to and fro; occasionally a stalk coils itself up into a spiral.
Fig. 1.
British Brachiopods (Terebratula and Crania).
The Brachiopoda, though presenting a certain outward resemblance to bivalved Mollusca, are quite distinct from this group. They are all marine, and all possess a bivalve shell. They grow attached to rocks (Fig. 1), usually by a horny peduncle or stalk passing between the two valves, or through a foramen in one of the latter; or, peduncle and foramen may be absent, one of the valves adhering by its surface to the rocks; some species of Lingula live in tubes in the sand or mud. They occur at all depths, from shallow water up to 2900 fathoms, but the largest number of species live at a depth of about 350 fathoms. Though found in all seas, the localities whence they have been obtained are comparatively few in number; but specimens are usually congregated in considerable numbers, in places where they do occur. The surviving species of Brachiopods constitute 75only a small remnant of a group that flourished abundantly in former epochs. There are about 150 recent, and over 6000 fossil species.
The Shell.—The valves of a Brachiopod shell differ from each other in size and shape, but each valve is in itself symmetrical, i.e., similar on each side of a middle line.
The valve through which the peduncle passes is termed the peduncle or ventral valve (Fig. 2, A), the other being the brachial or dorsal valve. The peduncle valve, which is usually the larger and uppermost, contains the bulk of the viscera; in the higher genera, calcareous bars or loops (Fig. 2, B) attached to the inner surface of the brachial valve form a support for the “arms” of the animal. The inner surface of the valves presents certain markings and depressions where the muscles have been attached (Fig. 6).
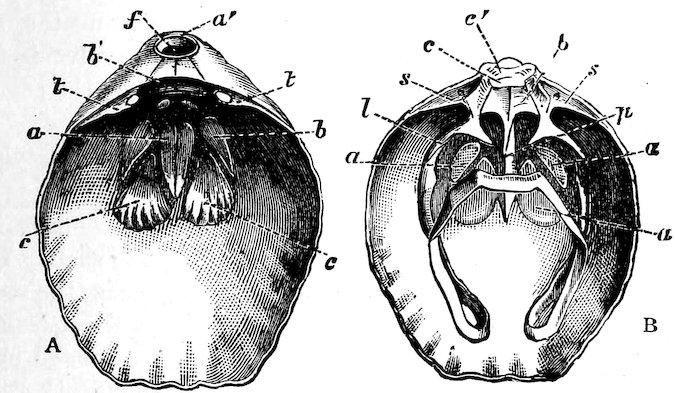
Fig. 2.
Magellania flavescens. Australia. Interior of valves.
A. Peduncle valve: f, foramen for peduncle, below which are the two small deltidial plates; t, hinge-teeth; a, b, c, muscle scars. B. Brachial valve, showing the reflected loop for support of the “arms.”
The shell is constructed of very minute prisms of calcareous substance imbedded in an organic matrix. In Lingula the shell is formed of alternating layers of horny and calcareous substance.
The shell-valves are either hingeless, or joined by a hinge in which teeth in the peduncle valve fit into sockets in the brachial valve. The Brachiopoda are primarily divided into two sections, Inarticulata and Articulata, based on the absence or presence of a hinge.
The division into Orders is based on the relation of the peduncle 76to the valves in its passage between them or through one of them. In the most primitive Brachiopoda (Lingulidæ), the peduncle simply passes out between the valves and not through a foramen or pore in one of them; hence the group is named Atremata (a, not, trema, pore). In the next group, including the families Discinidæ and Craniidæ, the peduncle passes through a fissure in the edge of the peduncle valve, the fissure in recent forms becoming closed round to form a slit-like foramen; this group is named Neotremata (neos, new, trema, pore). In the third group, Protremata (pro, in front of, trema, pore), which includes the Thecidiidæ, the peduncle lies at the apex of a triangular fissure in the peduncle valve, and secretes a calcareous plate to fill in the gap. In the fourth group, Telotremata (telos, final or complete, trema, pore), including the Terebratulidæ, etc., the triangular fissure in the peduncle valve is filled in by two calcareous plates termed deltidia, secreted by the edges of the mantle.
The valves are hingeless in the first two Orders (Inarticulata), and hinged in the last two (Articulata).
The Body.—The body usually occupies only a comparatively small space in the posterior or peduncle end of the shell. From each side of the body there is given off a thin expansion, the mantle which lines the inner surface of the shell. The space between the valves is termed the mantle-cavity. The mouth is situated in the centre of the front wall of the body or floor of the mantle-cavity. The front wall gives rise to a horseshoe-shaped platform surrounding the mouth and bearing on its upper edge ciliated tentacles, or cirri, which set up currents carrying food towards the mouth. In many genera the platform is produced into two coiled “arms” (Figs. 3 and 5), which fill up the mantle-cavity.
The name Brachiopoda (brachion, arm, pous, foot) was given to the group because these “arms” were supposed to be homologous with the Molluscan “foot.”
The mouth leads into a gullet, which opens into a stomach and intestine. In the more primitive forms the intestine terminates in a vent, but in the higher forms the distal end of the intestine has become atrophied, and consequently the gut ends blindly.
The body-cavity contains fluid, and is in communication with a system of sinuses in the lobes of the mantle (Fig. 5). Bands of muscles pass across from valve to valve. The peduncle consists of a horny outer sheath surrounding longitudinal and transverse bands of muscles.
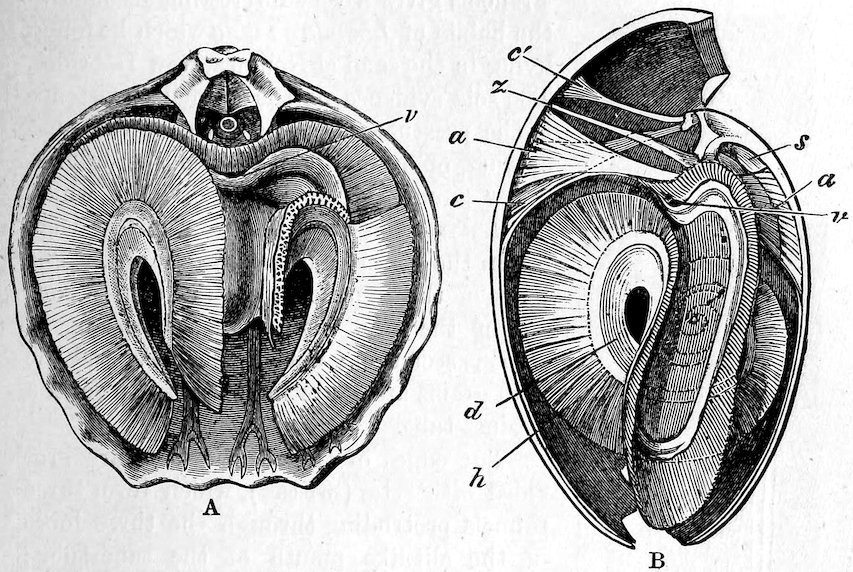
Fig. 3.
Magellania flavescens. (After Davidson.)
A. Interior of dorsal valve to show the “arms”; some of cirri removed on right side; v, mouth. B. Longitudinal section, with a portion of the animal.
The sexes are usually separate. The reproductive cells are formed in the body-cavity. The embryo swims freely for a short time before settling down and becoming fixed. The specimens exhibited in the case are arranged according to the following classification:—
| Section I. Inarticulata. | Order 1. Atremata. | Fam. Lingulidæ. |
| Order 2. Neotremata. | Fam. Discinidæ. | |
| Fam. Craniidæ. | ||
| Section II. Articulata. | Order 3. Protremata. | Fam. Thecidiidæ. |
| Order 4. Telotremata. | Fam. Rhynchonellidæ. | |
| Fam. Terebratulidæ. | ||
| Fam. Terebratellidæ. |
Order 1.—Atremata. Family Lingulidæ.—The Lingulas possess emerald green or golden brown duck-bill-shaped shells. Having no 78hinge, the dead valves of dried shells easily fall apart. The peduncle, which is sometimes over six inches in length, passes between the pointed posterior borders of the valves. Dr. François gives a very interesting account of the habits of Lingula anatina which he found living in the sand at Noumea, New Hebrides. The sole evidence of the animal’s existence is the presence, on the surface of the sand or mud, of a small, three-lobed slit (Fig. 4, upper figure). The tube (Fig. 4) in which the Lingula lives is about four inches deep, flat in the upper half, rounded below. The walls of the upper flat portion simply consist of the sand with a surface coating of mucous secretion; but in the lower end the sand grains are agglutinated so as to form a distinct tube.
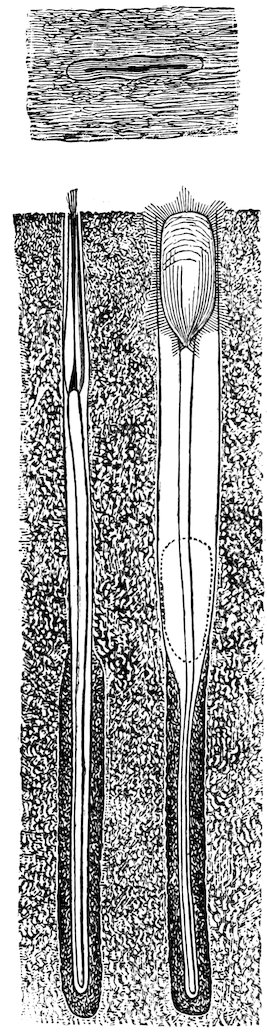
Fig. 4.
Lingula anatina in tubes in the sand; upper figure shows trilobed opening on surface of sand. Dotted line in lower figure indicates position in retraction. (After François.)
The edges of the mantle-folds are provided with setæ (bristles), which form three funnels protruding through the three lobes of the slit-like mouth of the sand-tube; currents enter by the lateral funnels and leave by the central.
On the least alarm the animal is rapidly withdrawn as far as the centre of the tube (see the dotted line of the shell in the figure), the surface slit and upper part of the tube being obliterated. Each of the arms forms a spiral with several coils (Fig. 5). The Lingulidæ are of exceptional interest, in that they furnish a very remarkable example of “persistence of type.”
Shells of Lingula occur in the earliest Palæozoic strata, and so closely resemble those of the present day, that often no difference can be observed either in the shape of the valves or in the muscular impressions on their inner surface (Fig. 6).
Lingula occurs in the Indo-Pacific, Australia, China, Japan, and the Pacific Islands. Glottidia, a smaller form, with two small curved plates on the brachial and a ridge on the peduncle valve, is found on the American coast of the Pacific, and in the Atlantic.
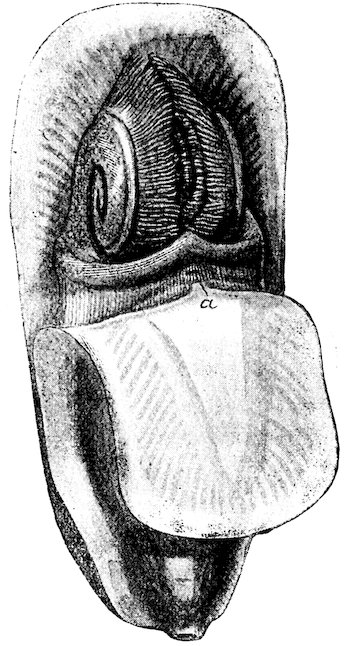
Fig. 5.
Lingula anatina, removed from shell, mantle reflected, coiled arms separated slightly; a, mouth. (Marginal setæ omitted.) Ventral aspect, three-quarter face.
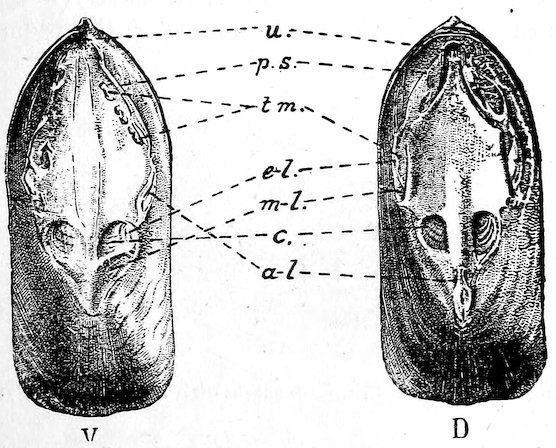
Fig. 16.
Lingula anatina. Interior of valves showing muscle scars.
V. Peduncle valve, D. Brachial valve.
80Order 2.—Neotremata. The Discinidæ includes two genera, Discina and Discinisca (Fig. 7), with orbicular conical shells, of horny calcareous composition; both valves are conical in the former genus, but in the latter the peduncle valve is flattened. Sometimes the embryos settle down on the parent shells, and we see a mass of shells in various stages of growth, as in the specimen of Discinisca lamellosa from Peru.
Fig. 7.
Discinisca lamellosa. Peru. (After G. Sowerby.)
A group of old and young specimens; largest showing foramen in peduncle valve, the rest showing brachial valves.
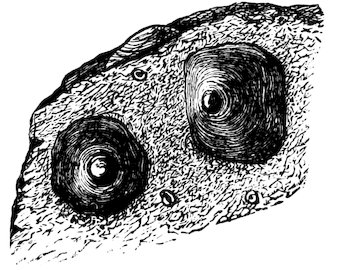
Fig. 8.
Three specimens of Crania anomala on a stone. Loch Fyne.
The Craniidæ form small limpet-like shells (Fig. 8) closely adherent to the rocks by the whole surface of the peduncle valve; although this valve is so named, no peduncle or foramen is found in this family. The Neotremata, like the Lingulidæ, are remarkable 81examples of persistence of type, since forms very similar to the present day Discinas and Cranias occur in the Palæozoic, Ordovician and Silurian strata.
A piece of rock, with several specimens of Crania anomala attached, is exhibited.
Order 3. Protremata.—This group, formerly very abundant, is now almost extinct, the Family Thecidiidæ representing the Order at the present day. Thecidium mediterraneum (Fig. 9) forms little oval boxes about a third of an inch in length, shaped somewhat like a pear cut in half (peduncle valve), and with a semicircular lid (brachial valve) working on a hinge on the upper flat surface. The foramen and peduncle are absent; but between the pointed end of the peduncle valve and the hinge is an area filled in by a calcareous plate characteristic of the Protremata.
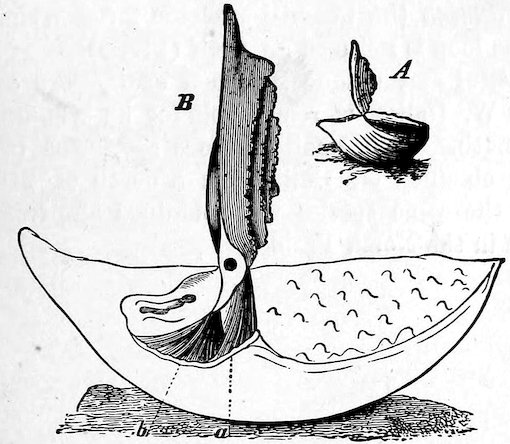
Fig. 9.
Thecidium mediterraneum. A, natural size. B, section through shell. Magnified.
The brachial valve opens like the lid of a snuff-box, and shuts down on the least alarm with the rapidity of lightning. The peduncle valve is fixed on the rocks by its convex surface. The species is common in the Mediterranean in from 30 to 300 fathoms, and is also found in the West Indies.
82Order 4. Telotremata.—This group, which at the present day contains the largest number of species, includes the Lamp shells, so called from their resemblance to an ancient lamp. The valves are joined by a well-marked hinge, the peduncle passes through the peduncle valve through a foramen completed by two plates secreted by the mantle edges, and the brachial valve has attached to it a calcareous scaffolding of processes or loops for the support of the “arms.”
The shells in this group are frequently ridged. Their colour is usually white, but sometimes red or yellow; deep-sea forms are generally vitreous.
Rhynchonella psittacea has a black shell with a pointed incurved beak; each of the arms forms a many coiled spiral and can be protruded beyond the shell; the brachial skeleton is comparatively small and simple, consisting of two separate processes.
In Terebratulina the brachial skeleton forms a simple loop; in Magellania the loop is reflected on itself (Fig. 2).
The beautiful and unique specimen of Dyscolia wyvillii, from 390 fathoms W. Indies, is remarkable for its size, being over two inches in length. The small vitreous specimens of Terebratula wyvillii were obtained off Chili from a depth of 2160 fathoms; specimens of the same species were obtained also from a depth of 2900 fathoms in the North Pacific.
The Tunicata are marine animals, the majority of which live, in their adult stage, a stationary life, fixed to the rocks or sea-bottom, but a comparatively small number are free-swimming.
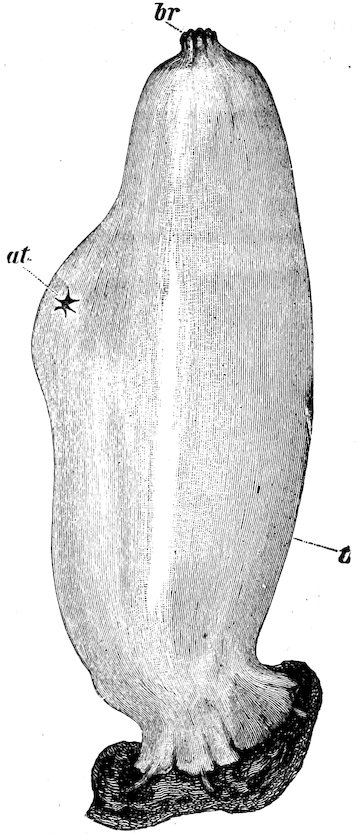
Fig. 1.
Ascidia mentula from the right side. at, atrial aperture; br, branchial aperture; t, test.
[After Herdman: Tunicata, Encyc. Britannica.]
They occur in the form of cartilaginous or leathery sacs, fleshy incrustations, solid fleshy masses, free-swimming, barrel-shaped animals, solitary or united into chains or hollow cylinders; or, lastly, of minute free-swimming tadpole-shaped organisms. To explain briefly the structure of a Tunicate, Ascidia mentula (Fig. 1), is selected. The 84animal, which lives on a muddy bottom, in from five to twenty fathoms, resembles a conical sac fixed by the broader end, of grayish green colour and about 4 inches in height. At the narrower end are two orifices, one terminal—the branchial orifice or mouth, and the other a little lower—the atrial orifice: the former has eight lobes and the latter six.
When the Ascidian is undisturbed, the orifices are wide open, and currents enter by the branchial and leave by the atrial orifice. On the least alarm, the orifices close, jets of water being at the same time squirted out; hence the popular name “Sea-squirts” given to these animals.
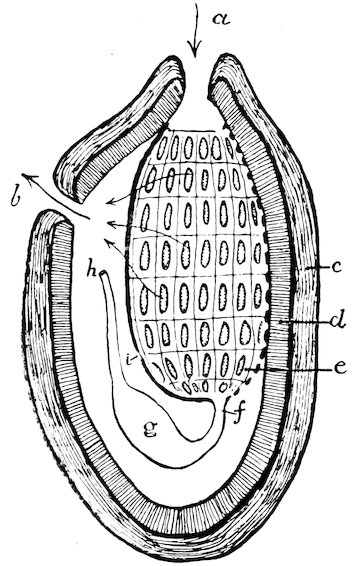
Fig. 2.
Diagrammatic section of Ascidia representing the three sacs, and the branchial sac as the pharynx or throat.
a, branchial; and b, atrial orifice; c, tunic or test; d, mantle; e, branchial sac; f, gullet; g, stomach; h, anal orifice; i, dorsal lamina; dotted line indicates the endostyle.
The Ascidian is orientated as follows: hold the animal with the branchial orifice pointing forwards and the atrial upwards; the branchial orifice will be anterior and the opposite end posterior; the atrial orifice will lie on the upper or dorsal aspect, the opposite aspect being lower or ventral, and the sides right and left. The aspects, in fact, correspond with those of a vertebrate animal. A 85vertical section roughly shows the animal to be formed of three concentric sacs (Figs. 2, 3). The outermost, which is tough and membranous, is called the Test or Tunic, the whole group owing its name to the presence of this protective covering.
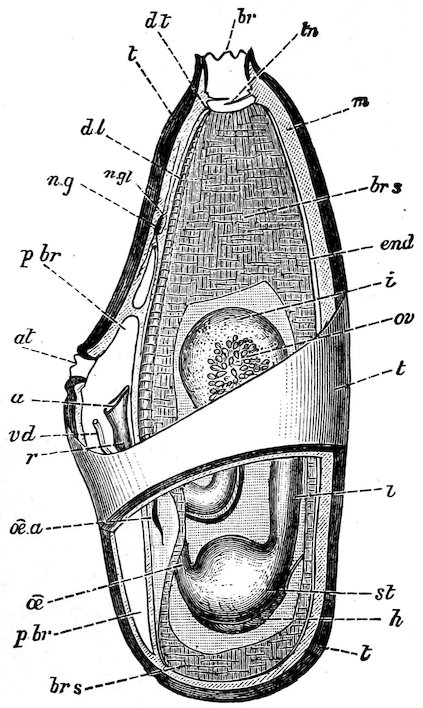
Fig. 3.
Diagrammatic dissection of A. mentula.
at, atrial orifice; br, branchial orifice; a, anal orifice; brs, branchial sac; dl, dorsal lamina; end, endostyle; m, mantle; ng, nerve ganglion; oea, orifice of gullet; pbr, peribranchial cavity; st, stomach; t, test; tn, tentacles.
(After Herdman: Tunicata, Encyc. Britannica.)
The middle sac, termed the Mantle, which almost corresponds in shape to the outer, is composed of connective tissue, muscle-fibres, blood-vessels, etc.; in spirit specimens, the mantle is shrunk away from the test except at the orifices and at a point behind, where vessels enter the test.
The innermost or Branchial Sac is attached behind the branchial orifice and along the ventral edge, but otherwise hangs free in the 86interior, the space around and outside of the sac being termed the atrial or peribranchial cavity.
The delicate walls of the branchial sac, which resemble fine muslin, are perforated by innumerable vertical slits, termed stigmata, arranged in transverse rows (Fig. 4).
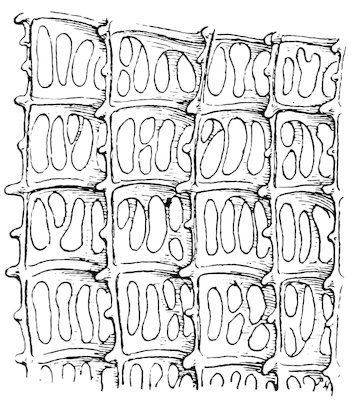
Fig. 4.
Ascidia mentula. Part of wall of branchial sac showing stigmata. Magnified.
The margins of the stigmata are lined with cilia which set up currents; and the water which enters by the branchial orifice, passes through the stigmata into the atrial cavity, and thence out through the atrial orifice. The walls of the branchial sac are chiefly composed of a sieve-like meshwork of fine blood-vessels arranged in transverse and longitudinal rows. The currents of water passing through the stigmata aërate the blood in the vessels. Besides the stigmata, the branchial sac has two relatively large orifices, viz., the branchial orifice or mouth, and, at the opposite end, the opening into the gullet. The branchial sac is, in fact, a capacious throat or pharynx (Diagram Fig. 2 and Fig. 14). Inside the branchial orifice is a circle of fine tentacles, which guard the entrance to the branchial sac. The food of the animal consists of minute animal and vegetable organisms.
It may be wondered how this food is secured, seeing that the currents of water are continually passing through the sieve-like walls of the branchial sac to the exterior again. Within the branchial orifice and above the branchial sac are two circular ciliated ridges with a groove between, which is full of viscid secretion; the cilia on the ridges direct particles into the groove where they are retained by the mucus.
87Passing backwards along the ventral edge of the branchial sac is a thick-lipped furrow, which appears like a rod in the thin-walled sac, and hence is called the endostyle. This organ secretes the mucus which is carried up by ciliary action to the circular groove in front of the branchial sac, and thence to the gullet along a fold or crest, termed the dorsal lamina, situated along the dorsal edge of the branchial sac.
The gullet opens into a large stomach situated posteriorly on the left side of the branchial sac. The stomach opens into the intestine, which, after forming a loop, terminates in the anal orifice or vent opening into the atrial cavity.
The tubular heart lies below the stomach, a remarkable feature in the circulation consisting in the periodic reversal of the blood current. An elongated nerve ganglion is situated between the branchial and atrial orifices.
Fig. 5.
Ascidian Tadpole with part only of the tail C. Magnified section.
N, nervous system with enlarged brain in front and narrow spinal cord behind n; N′, cavity of brain; O, the single cerebral eye lying in the brain; a, auditory organ; K, pharynx; d, intestines; o, rudiment of mouth; ch, notochord or primitive backbone.
(From Gegenbaur’s ‘Elements of Comparative Anatomy.’)
Ascidia mentula is hermaphrodite. The egg develops into a minute tadpole-like larva which swims about by means of its tail. Water entering by the mouth passes out through the gill-slits. A nerve-tube extending along the back and tail is swollen in front into a brain-vesicle; and underneath the long nerve-tube behind the brain is a stiff skeletal rod or axis—the notochord—which constitutes the rudiment of a backbone. Inside the brain are two unpaired sense organs, an eye and an organ of hearing (Fig. 5). After swimming freely for a few hours, the larva settles down head foremost and 88fixes itself by papillæ on the anterior end (Figs. 6, 7). Presently the tail becomes absorbed, and the posterior end of the nerve-tube, and the brain with its eye and hearing organ, undergo atrophy, the nerve ganglion of the adult alone representing the cerebrospinal axis of the larva. The branchial sac and intestines develop greatly, and growth proceeds in such a manner that the mouth is pushed round to a position opposite to the fixed area, and gradually the animal becomes the adult ascidian.
Fig. 6.
Degeneration of Ascidian Tadpole to form the adult. The black pieces represent the rock or stone to which the Tadpole has fixed its head.
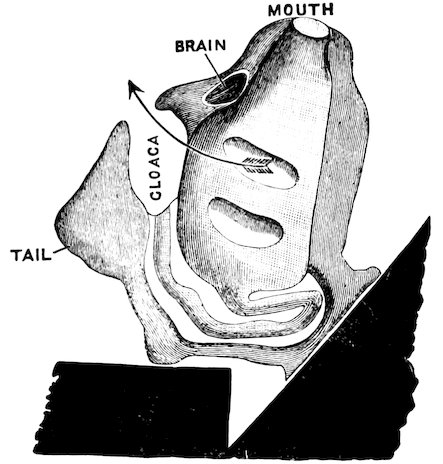
Fig. 7.
Very young Ascidian with only two gill-slits.
(Figs. 6, 7, from Lankester’s ‘Degeneration.’)
89This wonderful metamorphosis presents a striking example of DEGENERATION resulting from the adoption of a fixed mode of life. The active free-swimming larva with its brain, eye, hearing organ, and muscular tail becomes transformed into a comparatively inert sac.
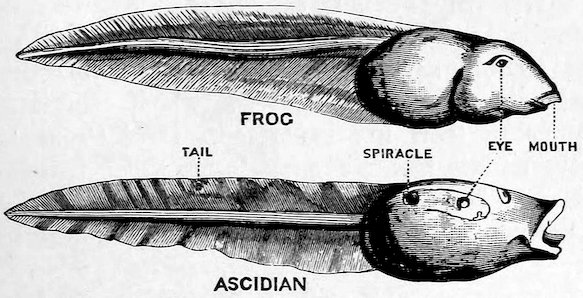
Fig. 8.
Tadpole of Frog and Ascidian. Surface view. (Lankester’s ‘Degeneration.’)
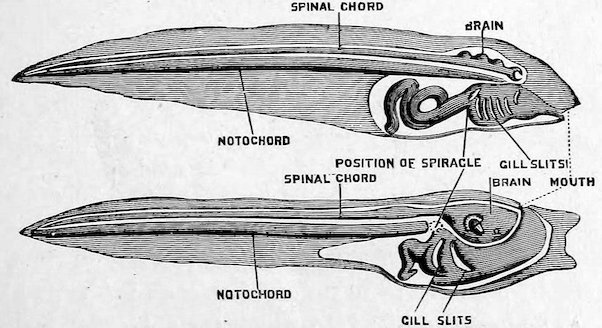
Fig. 9.
Tadpole of Frog and Ascidian. Diagram representing the chief internal organs. (Lankester’s ‘Degeneration. A chapter in Darwinism.’)
The tadpole of an Ascidian resembles that of a frog (Figs. 8, 9), not merely superficially, but also in its general structure and mode of development. The Tunicata are now generally regarded as a degenerate offshoot from the ancestral stock of the Vertebrata, in that the larva possesses a skeletal rod (rudimentary backbone) separating the dorsally situated nerve-tube (cerebrospinal axis) from the ventrally situated intestinal tube, the existence of the cerebral 90eye in the Ascidian tadpole further tending to confirm the truth of this theory. Apart from a knowledge of the course of their development, Tunicata would have been classed among the Invertebrata, but the structure of the larva clearly reveals the affinities of the group to the backboned animals.
Ascidia mentula belongs to the group of Simple Ascidians which are all fixed, and are either solitary or joined into colonies in which each individual or ascidiozooid has a distinct test of its own. In the Compound Ascidians, which form colonies by budding, the ascidiozooids are buried in a common investing mass and have no separate tests. In a third group, the Salpa-like Ascidians, the ascidiozooids are united to form free-swimming colonies shaped like hollow cylinders open at one end. The above three groups belong to one great Order—the Ascidiacea. A second Order, Thaliacea, includes the free-swimming Salpa and Doliolum, which exhibit alternation of generations in their life history. A third Order Larvacea, includes very minute free-swimming forms which possess a tail in the adult stage. There are sixteen families of Tunicata.
The following is a tabular view of Prof. Herdman’s classifications:—
| Order I. Ascidiacea. | Sub-order 1. | Ascidiæ | Simplices, 4 Families. |
| Sub-order 2. | „ | Compositæ, 7 Families. | |
| Sub-order 3. | „ | Salpiformes, 1 Family. | |
| Order II. Thaliacea | 3 Families. | ||
| Order III. Larvacea | 1 Family. | ||
The Ascidiacea include the great majority of species. With the exception of the one genus Pyrosoma, they lead a fixed or stationary life.
The Simple Ascidians are mostly solitary; in a few forms, however, colonies arise by budding from stolons, but each individual has a distinct test. The four families into which the sub-order is divided are chiefly characterised by the nature of the test, the number of lobes round the branchial and atrial orifice, and the character of the branchial sac.
In the family Molgulidæ the tough membranous test is often coated with sand; the branchial aperture is six-lobed, the atrial four-lobed, the branchial sac has long folds or pleats, and the stigmata are curved or arranged in spirals.
91Molgula gigantea, which is one of the largest of the Ascidians, and which attains a length of over thirteen inches, forms a tough conical sac; the branchial and atrial orifices at the upper end have six and four lobes respectively. The test is leathery, smooth above, but coated with sand below. The exhibited specimen, which comes from the Straits of Magellan, has several specimens of the stalked Boltenia legumen attached to the lower part of the test.
The curious Molgula oculata (Fig. 10) has a soft oval or rounded body coated with sand. The branchial and atrial orifices have respectively six and four lobes. Specimens grow attached to the rocks and also live free in the sand. The surface of the test is provided with hairs, which adhere to the rocks and collect particles of sand. The adhesion not being very firm, specimens are easily detached by currents and collected into heaps by the eddies; when living in the sand only the two dark orifices are visible. The sand coating has been supposed to confer protection by mimicry of the environment; but Professor Lacaze Duthiers found, much to his chagrin, that the sandy tests of his specimens were of no avail in securing them from being devoured by crabs who seemed to scent their prey from afar.
Fig. 10.
Mogula oculata.
a, branchial; b, atrial orifice.
In the family Cynthiidæ the test is usually leathery, the branchial and atrial apertures four-lobed, and the branchial sac folded into longitudinal pleats.
92The genera Boltenia and Culeolus include species in which the body is attached to a peduncle.
The large exhibited specimen of Boltenia pachydermatina is 28 inches in length, the head being 4 and the stalk 24 inches long. The two four-lobed apertures are along one edge, the branchial being the lower; the body is marked with long deep furrows, and the stalk with transverse wrinkles. Culeolus perlucidus, from 1600 fathoms in the Southern Ocean, is in the form of a small pear-shaped head on a slender stalk, the total length being 4½ inches. The branchial orifice forms a transverse slit with raised lips near the stalk, the slit-like atrial orifice being near the rounded end of the body. Culeolus moseleyi, another slender-stalked form, was obtained from 2425 fathoms in the Central Pacific.
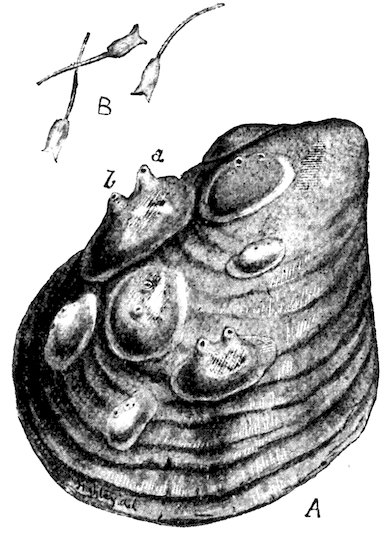
Fig. 11.
A. Styelopsis grossularia on shell. B. Tadpoles of same, × 9. a, branchial; b, atrial orifice. (B, after Sir J. Dalyell.)
The little Cynthiid Styelopsis grossularia (Fig. 11), popularly known as the “Currant Squirter,” occurs in the form of bright red hemispherical blobs on stones and shells; when undisturbed, the branchial and atrial orifices expand and project upwards. The eggs are brilliant red in colour. Sir John Dalyell was the first to discover the tadpole form, which is about ⅒ inch long (Fig. 11, B), and to 93observe the tadpoles become fixed and develop into fixed Ascidians. He calls the active little swimming larvæ “Spinulæ,” from their resemblance to small pins.
The family Ascidiidæ includes forms with a gelatinous or cartilaginous test; the branchial and atrial orifices usually have 8 and 6 lobes respectively; the branchial sac is without folds.
Ascidia mentula, described above, belongs to this family.
Chelyosoma is characterised by the test forming tortoise-like horny plates on the upper surface. The exhibited specimen of C. macleayanum (Fig. 12) comes from Greenland; the upper hemispherical part of the test is divided into 8 plates; the branchial and atrial orifices are situated in the joints between the plates.
Fig. 12.
Chelyosoma macleayanum, slightly enlarged. a, branchial; b, atrial orifice.
The fine specimen of Phallusia mammillata from Naples consists of several individuals partly fused together; the branchial and atrial orifices are wide open, and the mantle can be seen through the thick knobby translucent test.
In Rhodosoma the test is modified so as to form stiff plates recalling the valves of a bivalve shell. One plate is attached to the rocks, the other closing against the first like a lid; the anterior end of the animal with its branchial and atrial orifices is visible only when the 94lid is open. The Mediterranean species R. callense (Fig. 13) grows attached to the rocks. The little exhibited specimen is on a fragment of shell in front of a black patch. The figure shows specimens with the lid open and closed.

Fig. 13.
Rhodosoma callense, × 10. A, “valve” open; B, shut. a, branchial; b, atrial orifice. (After Lacaze Duthiers.)
Family Clavelinidæ. The body is attached to a creeping stolon or mass of stolons, from which new individuals arise by budding. The other three families of Simple Ascidians included solitary forms, but the Clavelinidæ are social, and form colonies wherein each individual has its own test.
Clavelina lepadiformis (Fig. 14) forms graceful crystal vases about an inch in height. The figure shows one individual, but usually the processes at the base extend out as stolons whence other individuals arise.
Diazona violacea, from Cornwall, forms beautiful purple disk-shaped colonies in which the ascidiozooids arise from a basal mass of stolons. Sometimes the ascidiozooids die down, leaving only a smooth violet pad, which in due time produces a new crop of ascidiozooids.
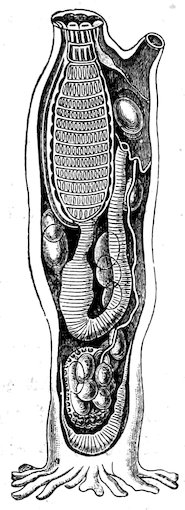
Fig. 14.
Clavelina lepadiformis; diagrammatic, showing the anatomy. The oval bodies are the eggs; at lower end lies the tubular heart; the root-like processes at the base grow into stolons, whence other ascidiozooids arise.
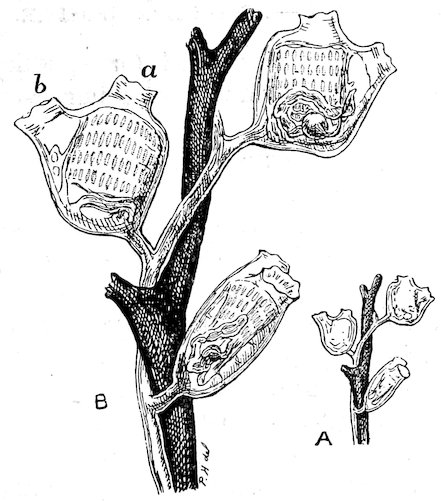
Fig. 15.
Perophora listeri; A, slightly, B, further magnified. Ascidiozooids in right, left, and lateral aspects.
a, branchial; b, atrial orifice.
96The remarkable Rhopalæa neapolitana, from Naples, may be roughly compared to an hour-glass with a very long constriction. The test is smooth in the upper part, but knobby and encrusted with foreign bodies below. The upper or thoracic end contains the branchial sac, and the lower or abdominal portion the stomach, heart, and reproductive organs, the gullet and intestine traversing the whole length of the narrow central region. Although from its general structure Rhopalæa is a Clavelinid, it is not certainly known to produce buds.
Perophora listeri (Fig. 15) occurs in the form of little jelly-like transparent blobs rising by short stalks from a silvery thread-like stolon. Owing to their small size and transparency, it is possible to examine specimens alive under the microscope, the currents passing through the stigmata in the walls of the branchial sac, and the beating of the heart being distinctly visible. The rapid motion of the cilia surrounding stigmata gives the appearance of dark wheels all rotating in the same direction. The heart beats so as to drive the blood current so many times in one direction, and then after a short pause, in the reverse direction.
The exhibited specimen growing on an oyster shell, is from Plymouth.
The Compound Ascidians are fixed forms, which give rise to colonies by budding, the individuals being immersed in a common mass and not possessing separate tests.
Although reduced to an extremely small size each individual or ascidiozooid of a colony possesses the same organs as a large Simple Ascidian, excepting that the former does not possess a separate test. Frequently the individuals of a colony are grouped into systems, in which the atrial orifices open into a common cloaca. The little ascidiozooids vary greatly in shape in the different families. In the Polyclinidæ, for instance, they are long, the organs being so to speak, drawn out, and being arranged in three regions, the thoracic, abdominal and post-abdominal, the first region containing the branchial sac, the second the stomach, and the third the heart and reproductive organs. In the Distomidæ, the body exhibits two regions, thoracic and abdominal, the heart and reproductive organs lying alongside of the stomach. The Botryllidæ comprise only one region, the stomach and the other organs being situated by the side of the branchial sac.
97The Compound Ascidians include seven families which are characterised chiefly by the method of bud formation, and by the arrangement of the organs into one, two, or three regions.
It is only possible, from limits of space, to refer to a few interesting forms.
The species of Botryllus are those most commonly met with. They form richly coloured gelatinous incrustations on rocks and seaweeds. B. violaceus (Figs. 16, 17, and 18 D) is blue with white lines; B. smaragdus, green; B. marionis, brown with white and carmine; B. castaneus, purple, and so on. The individuals are arranged in circular systems with the branchial orifices round the circumference and the atrial orifices opening into a common central cavity (Fig. 17), the whole colony being composed of groups of systems.
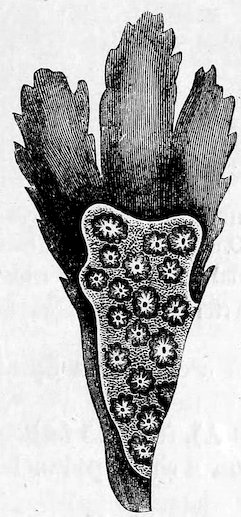
Fig. 16.
Botryllus violaceus on seaweed. (After H. Milne-Edwards.)
The exhibited specimen of B. violaceus was grown in the tanks of the Biological Station at Plymouth. The red specimen of B. aurolineatus, from Naples, shows well the branchial and cloacal orifices. In Botrylloides, the individuals form elliptical or elongated systems.
Colella thomsoni was obtained near the Philippines at a depth of 10 fathoms. The specimen, which is about 7 inches in length, 98resembles an elongated head of clover on a thickened stalk. The individuals which compose the head are arranged in spiral lines, the atrial orifice of each ascidiozooid opening separately and not into a common cloaca.
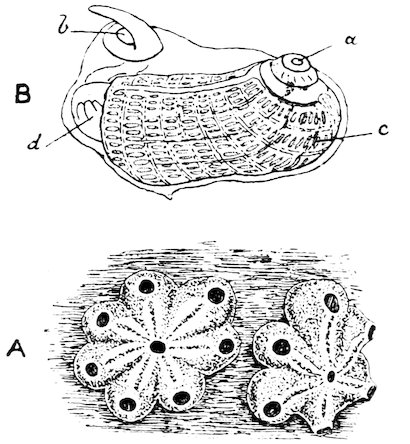
Fig. 17.
A. Botryllus violaceus, magnified, showing two systems of 6 and 7 ascidiozooids. B. One ascidiozooid extracted.
a, branchial; b, atrial orifices; c, branchial sac; d, stomach.
(After H. Milne-Edwards.)
Colella quoyi (Fig. 18 A), from 25 fathoms off Kerguelen Island, forms a rounded head on a short peduncle, the total height being one inch. The ascidiozooids are arranged in vertical lines in the “head,” each line consisting of a double zigzag series.
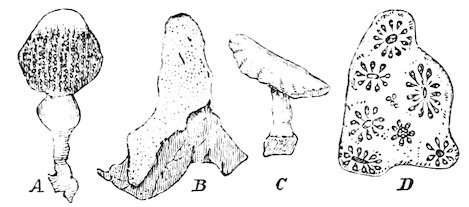
Fig. 18.
Colonies of Ascidiæ compositæ, natural size. A. Colella quoyi. B. Leptoclinum neglectum. C. Pharyngodictyon mirabile. D. Botryllus.
(After Herdman, Challenger Report and Encyclopædia Britannica.)
Julinia ignota, from the Antarctic regions, forms long narrow 99colonies, which attain a length of nearly three feet. One end is attached, the rest of the colony apparently lying along the sea-bottom.
Amaroucium roseum from Naples forms translucent gelatinous masses; a slice is exhibited, showing the long slender ascidiozooids immersed in the mass.

Fig. 19.
Pyrosoma elegans, natural size. A. Side view of entire colony. B. End view of open extremity.
(Herdman: Tunicata, Encyclopædia Britannica.)
Pharyngodictyon mirabile (Fig. 18 C), from 1600 fathoms in the Southern Indian Ocean, resembles a small mushroom, and is about one inch in height. This species is one of the few deep-sea Compound Ascidians.
Leptoclinum albidum is a common and widely distributed species; it occurs in the form of thin white crusts. The glistening white appearance is due to the common test being densely crowded with minute stellate spicules of carbonate of lime.
The specimen of Leptoclinum neglectum (Fig. 18 B) encrusts a fragment of sponge.
Goodsiria pedunculata from the Straits of Magellan, forms a rounded cartilaginous mass attached by a short peduncle; sometimes several masses are attached to each other. Each of the small dark oval areas on the surface corresponds to the branchial and atrial orifices of one ascidiozooid.
The Salpiform Ascidians comprise only one genus, Pyrosoma, which occurs in the form of free-swimming colonies shaped like hollow cylinders closed and rounded at one end and open and truncate at the other (Fig. 19). The wall of the cylinder is formed of a single layer of ascidiozooids (Fig. 20), so arranged that all the atrial orifices open into the interior of the cylinder, and all the branchial orifices on the exterior, the two kinds of orifices being at opposite ends of the body, and not close together, as in most simple and compound Ascidians.
100Specimens vary in size from a few inches to upwards of four feet in length, and, as the name of the genus implies,[22] they are brilliantly phosphorescent. Sometimes they occur in innumerable multitudes, giving rise to a zone of greenish light extending for miles. Professor Moseley records that during the voyage of the Challenger in the North Atlantic a huge specimen of Pyrosoma spinosum, four feet in length, was captured. On tracing his name on its body, the word came out in letters of fire.
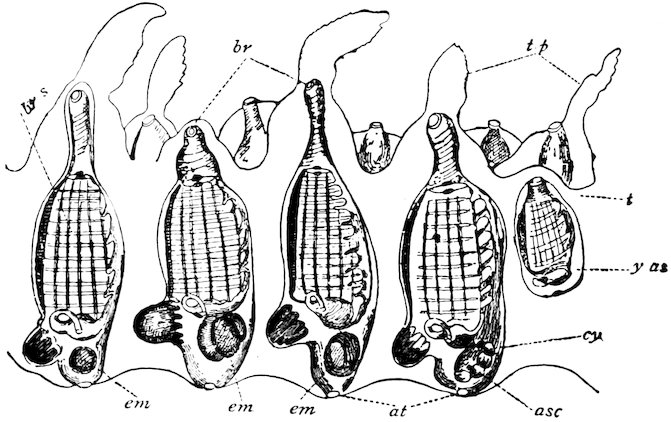
Fig. 20.
Section through wall of Pyrosoma, magnified, showing a single layer of ascidiozooids.
br, branchial; at, atrial orifice; tp, process of the test; br s, branchial sac.
(Herdman: Tunicata, Encyclopædia Britannica.)
In Pyrosoma elegans (exhibited), from Naples, the ascidiozooids are arranged in verticils, and the mouth of the cylinder is surrounded by a movable diaphragm; the outer end of each ascidiozooid is provided with a membranous spine. Six species of Pyrosoma are known. Pyrosoma atlanticum is found in the tropical Atlantic and Antarctic; P. giganteum in the Atlantic, Pacific, and Antarctic; and P. spinosum in the South Atlantic.
The Thaliacea are free-swimming Tunicates, which exhibit alternation of generations in their life history. There are three families, Salpidæ, Octacnemidæ, and Doliolidæ.
Salpidæ.—The Salpas are transparent barrel-shaped organisms 101which occur in abundance at the ocean surface. They are so transparent that they are rarely seen, except in calm weather from the side of small boats; yet they frequently swarm in countless multitudes. From five to ten bands of muscles partially or entirely surround the body, like hoops. The branchial and atrial openings are at or near the opposite ends of the body. The branchial sac has almost disappeared, the dorsal lamina and ventral gutter (or endostyle) alone remaining, the interval between the two on each side representing an enormous stigma; the dorsal lamina, or “gill” is the transversely striated band passing obliquely across the body and forming the only barrier between the branchial and atrial cavities. Water enters at the mouth, and, by the contraction of the muscle-hoops, is driven out through the atrial aperture at the opposite end, which is then closed by a sphincter muscle. The elastic walls of the body expand, and water again enters through the mouth, the valve-like lips of which prevent its being driven out that way. The Salpa swims along in jerks, and along with each gulp of water takes in Radiolaria, Foraminifera, etc., which are retained by the mucus of the endostyle and carried to the gullet. The Salpa, in fact, lives, as Professor Brooks observes, in a “living broth,” so abundant is the food supply.
The intestines usually form an oval mass termed the “nucleus,” which is a conspicuous object at the posterior end.
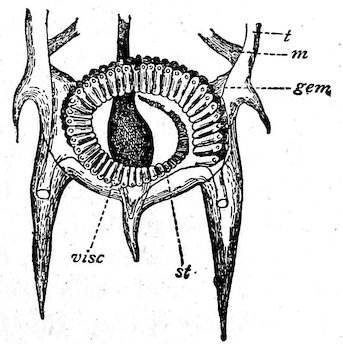
Fig. 21.
Posterior part of solitary form of Salpa democratica-mucronata, showing a chain of embryos nearly ready to be set free.
gem, young chain of Salpæ; st, stolon; t, test; visc, visceral mass.
The solitary Salpa above described is asexual. In the ventral region of its body it forms a stolon which becomes segmented into a series of buds (Fig. 21). As the stolon grows the end series of buds 102breaks off in the form of a chain and swims away, other chains being detached in succession. A chain is formed of individuals arranged in two rows, the individuals in each row being alternate (not opposite).
Each individual of a chain differs from the solitary individual in shape, arrangement of muscle bands, etc., but especially in having reproductive organs. The chain Salpid is hermaphrodite; the embryo develops into a solitary asexual Salpa which produces the chains by budding. The wonderful life history of Salpa was discovered by the poet Chamisso during a voyage round the world in 1819. He observes: “A Salpa mother is not like its daughter or its own mother, but resembles its sister, its granddaughter, and its grandmother.” Here we have an example of “alternation of generations,” a sexual generation (chain form) giving rise to an asexual generation (solitary form), which latter produces the sexual generation.[23]
Most of the species of Salpa have double names owing to the chain and solitary forms having been regarded as distinct species before they were known to be phases in the life history of one and the same species. Salpa runcinata-fusiformis, solitary form (Fig. 22 B), is barrel-shaped, truncated at each end, with terminal orifices, and with nine muscle-bands on the dorsal surface, some of which converge towards each other. An individual of a chain (Fig. 22 A) is fusiform, with six muscle-bands, and with the orifices not terminal, but at each end of the dorsal surface.
The solitary form of S. africana-maxima is barrel-shaped, with truncated ends and terminal orifices, and with nine broad parallel muscle-bands. The chain form is conical at one end, with six bands, and with orifices on the dorsal surface. The exhibited specimen of the chain form, which is in an early stage of growth, contains 202 individuals. The solitary and chain individuals of Salpa costata-tilesii attain a length of six to eight inches. The solitary form has eighteen muscle-bands and two large spines at the posterior end. The individual of the chain has five muscle-bands. A chain of three individuals is exhibited.
Salpa pinnata produces a circular chain; the exhibited specimen of the solitary form shows a small chain about to be detached; a circular chain of six individuals is also exhibited. Species of Salpa abound in all seas, but specimens from Naples have alone been exhibited on account of their good preservation.
103Family Octacnemidæ includes O. bythius, a deep-sea Salpid, in which the body forms a flattened disk produced into eight radiating lobes.
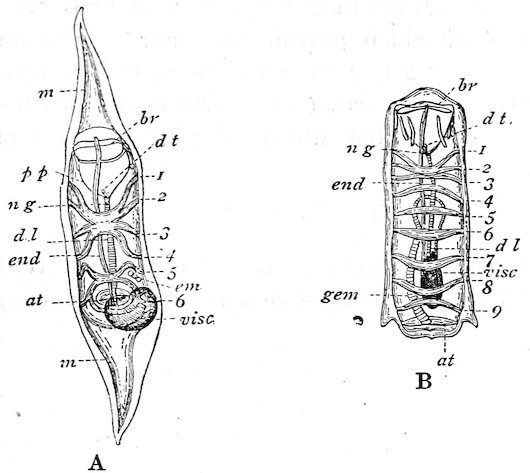
Fig. 22.
Salpa runcinata-fusiformis. A. Chain form. B. Solitary form. 1–9, muscle bands; em, embryo; m, mantle; visc, visceral mass or nucleus.
(Herdman: Tunicata, Encyclopædia Britannica.)
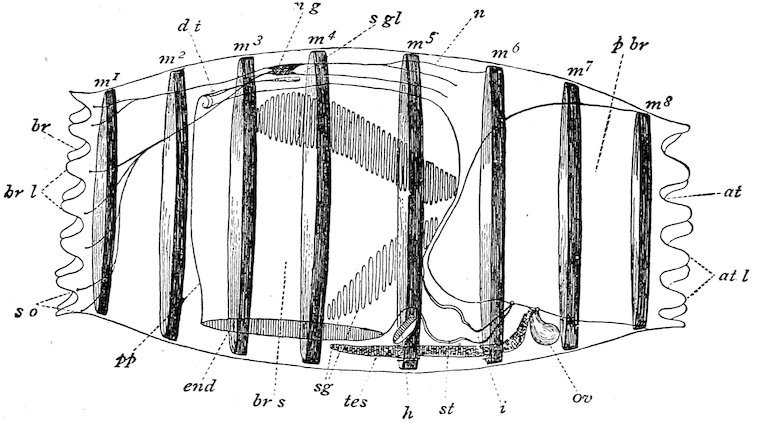
Fig. 23.
Doliolum denticulatum, sexual generation, from the left side. m1-m8 muscle bands; at, atrial; br, branchial apertures; br s, branchial sac; sg, stigmata; st, stomach; ng, nerve ganglion; so, sense organs.
(After Herdman, Encyclopædia Britannica.)
Family Doliolidæ. The body is cask-shaped and surrounded by circular hoops. The branchial and atrial orifices are at the opposite 104ends. The branchial sac is pierced by two oblique bands of stigmata (Fig. 23 sg). The life history is very complicated. The egg develops into a tailed larva, which develops into a “nurse”; the latter is asexual, and produces three kinds of buds on a stolon, viz. (1) nutritive buds which provide the “nurse” with food, (2) foster forms which are set free as cask-shaped bodies with eight broad muscle-bands, and (3) sexual forms which are attached for a time to the foster forms, but which later become free and give rise to the egg.
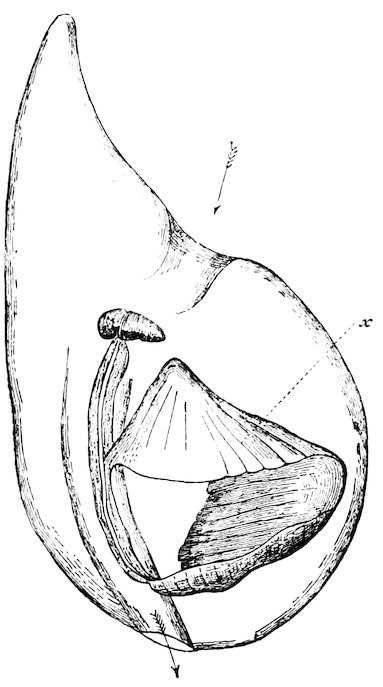
Fig. 24.
Oikopleura cophocerca in its “house” (after Fol); seen from right side, × 6. Arrows indicate course of the water; x, lateral reticulated parts of the “house.”
The Larvacea are very minute Tunicata which live at the surface and swim by means of a tail-like appendage, resembling in this and certain other respects the tadpole larva of other Tunicata. They are able to form a temporary test or “house” many times larger than the body (Fig. 24). The organism itself, which is almost lost 105in its large test, is the little hammer-shaped body in the centre of the figure; the streaked areas bound a space in which the tail lashes vigorously. The animal can leave its test and secrete another in a few hours.
The tail is attached to the under or ventral surface of the tiny little barrel-shaped body, and usually points forwards; a skeletal rod, the urochord, runs along its length. The branchial sac has two ciliated openings or gill-clefts leading directly to the exterior, and not opening like the stigmata of the other orders into an atrial cavity.
The order contains one family, the Appendiculariidæ, and four genera, and is represented in all seas.
Oikopleura cophocerca, one of the largest forms, is about half an inch in length. The exhibited specimens came from St. Andrews, Fife. Professor McIntosh reports that occasionally specimens of this species occur in immense quantities, the tow-nets being filled with them.
In the Starfish Gallery is exhibited a series of the animals belonging to the class Echinoderma; of these the Starfishes are the best known, while others are the Sea-Lilies, Sea-Urchins, and Sea-Cucumbers or Sea-Slugs.
A small collection of various kinds of Worms is also exhibited in this Gallery (Wall-cases I.–III.).
Six table-cases contain the dried Echinoderms arranged in systematic order. The seventh is devoted to preparations, models, and figures illustrative of the structure and life history of various members of the group.
An inspection of that Case and the accompanying woodcuts will make clear the distinctive characters of the Echinoderma. Unlike that of a Crayfish or a Mussel, the body does not appear to be divided into two equal or symmetrical halves, though it really is; this is due to the possession of a number of rays, of which there are ordinarily five. The skin is strengthened by the deposition in it of carbonate of lime, which may be in the form of continuous plates or bars, or of separate scattered spicules. A series of tube-feet or suckers (podia) are generally developed along each ray, and these are supplied by a system of water-vessels peculiar to Starfish and their allies. These rays are often called “ambulacra.”
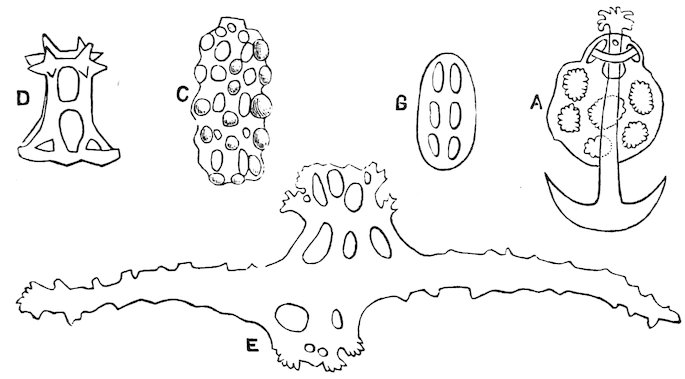
Fig. 1.
A. Anchor and plate of Synapta. B, C. Tables of Holothuria impatiens; and D. Holothuria atra: from various aspects. E. Spicule from sucker of Stichopus variegatus, magnified about 200 times.
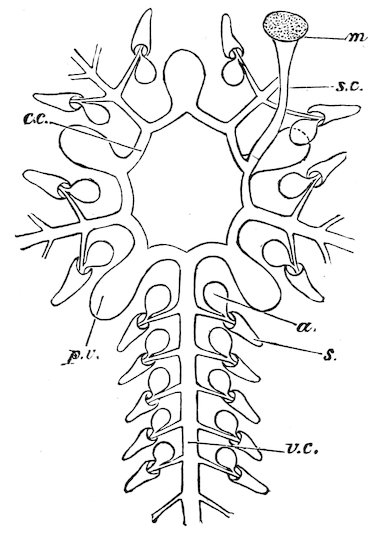
Fig. 2.
Diagram of Water-vessels.
c.c. Circular canal, with p.v., its Polian vesicles; from it a radial canal (v.c.) is given off along the lower surface of each arm; this supplies, by side branches, the suckers, s; connected with each sucker is a contractile swelling or ampulla (a). The circular canal is in connection with the exterior by s.c., the stone-canal, and opens to it by the madreporite (m).
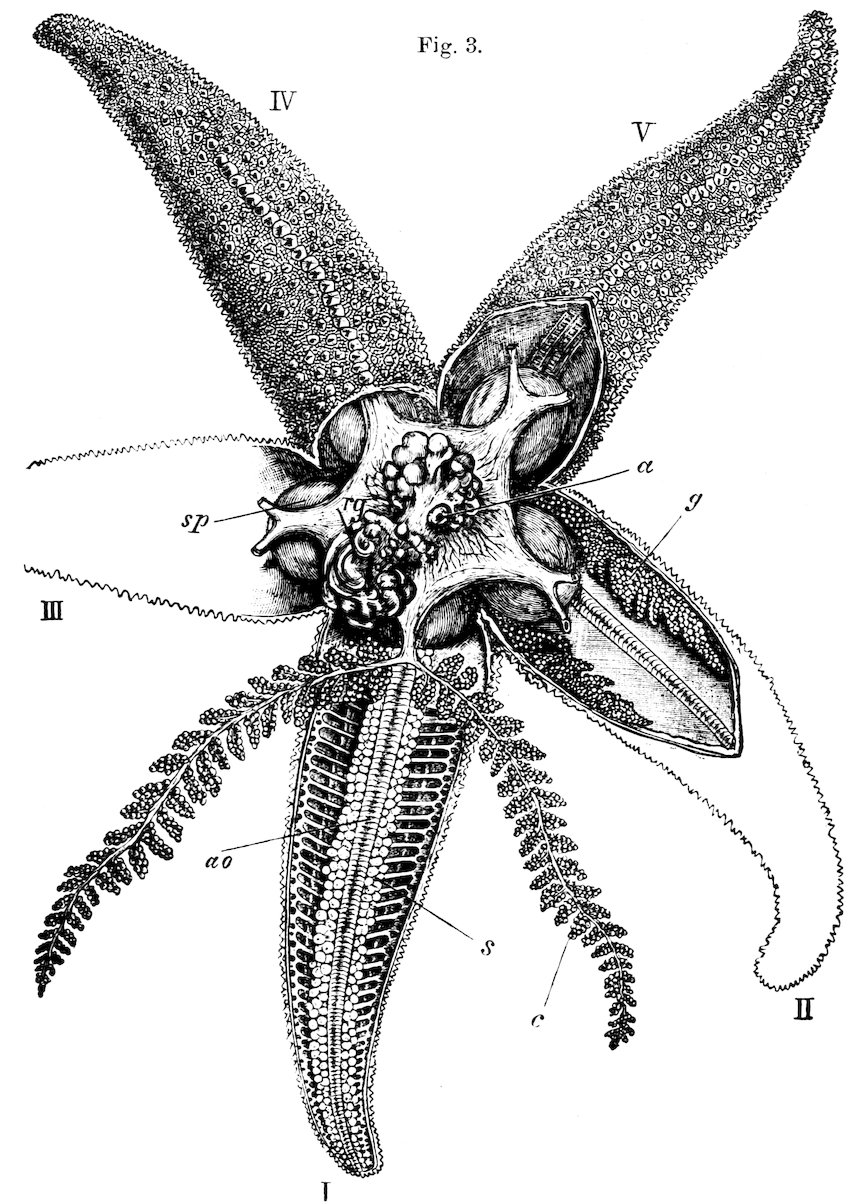
Fig. 3.
Figure of a Starfish (Asterias rubens).
In the ray marked I. the skin has been removed from the upper surface, and the ambulacral ossicles (ao) and the podia (s) are seen in situ; the blind outgrowths (c) from the central stomach (sp) have been dissected out. In II. the gonads (g) are exposed; and in the centre above the stomach the rectal glands (rg) are to be seen. The anus (a) is seen to be subcentral in position.
109In the body of the Starfish (Fig. 3) the arms are seen to be continuous with the disk and to contain portions or prolongations of the chief organs. The middle of the arm is occupied by two rows of hard pieces (ambulacral ossicles), the fellows of which make an open angle with each other, and so form an open ambulacral groove; along this we find the suckers, the water-canal that supplies them, the blood-vessel of the arm, and a nerve-cord. At the centre of the disk is the mouth. The ossicles at the sides of the arms bear spines, which vary in different species; the surface of the back is supported by a network of hard pieces, and through the intervening spaces there project membranous pouches, which are respiratory in function. The modified plate on the upper surface opens into a tube by means of which the water-vessels communicate with the exterior; this plate is known as the madreporite (Fig. 2, m).
The organs for masticating the food are most highly developed in the regular Echinoids, where the complex apparatus known as the “Lantern of Aristotle” is found (Case 38) to consist of five sets of pieces; the tooth is strong and bevelled at its free end; it is supported by triangular jaws on either side, a pair uniting and having the form of an inverted pyramid; these alveoli are connected with their neighbours by oblong pieces (falces); above these there are elongated bars, which are hinged on to the inner end of the falces and have their outer ends free. The whole lantern is connected to the test by muscles which pass from its sides to the auricles or upstanding pillars which lie round the mouth; and, owing to this muscular apparatus, the teeth are capable of complicated and various movements.
In the Ophiuroids the edges of the mouth-slits are provided with short spinous processes, varying a good deal in arrangement, but never having, apparently, any other function than that of a filtering-apparatus; in the Starfishes the plates round the mouth have a supporting function only; in Crinoids and Holothurians the mouth is unarmed; the latter are often remarkable for a deposit of calcareous plates in the walls of the gullet, and in the former the grooves on the arms are the lines along which food comes to the mouth.
Echinoids live on seaweeds and the animals that are found on them; such as have no teeth, like Spatangus (Case 32), use their spout-like mouth to take up the sand and débris on which they move, and from which they extract some nutriment. Ophiuroids live on the smaller foraminifera; Asteroids on dead fishes (as line-fishermen well know), oysters, and other molluscs, and even on specimens of their own particular species; Holothurians on shell or coral débris and the minute organisms it contains; and Crinoids on small tests of foraminifera and on the adults of small and larvæ of larger crustacea.
110In a number of Echinoids and Asteroids some of the spines are specially modified to act as seizing-organs—the free end being divided into two, three, or rarely four pieces, which are moved on one another by special muscles. These minute organs were regarded by earlier observers as parasites, and were named pedicellariæ; they may be movable, when they have a stalk, or the stalk may be absent and the valves sessile. Considerable difficulty attaches to the determination of the use that these organs may be to their possessors; but there is reason to suppose that they may act as cleansing-organs by removing minute particles of dirt, and as temporary organs of fixation, while M. Prouho has observed their use as organs of defence.
Echinoderms move but little; the unstalked Crinoids, if they cannot find stones or worm-tubes around which to attach themselves, swim by beating the water with their delicate arms, five being raised and five depressed alternately. The Echinoid or Asteroid is able to move by the aid of its podia or so-called ambulacral feet, which become erected by being filled with water, and are then contracted; by means of this contraction movement is effected; a similar kind of locomotion obtains with the pedate Holothurians; in the Ophiuroids the flexible arms either serve as the organs of movement, or act as an apparatus whereby the creature becomes coiled round the branches of corals (see Case 20).
Echinoderms are often of exceedingly bright colours, as is shown by the pictures on the wall, and are very conspicuous objects; this may, apparently, be associated with disagreeable tastes or odours; sometimes they cover themselves over with seaweed, and so hide their brilliancy; the spines of some forms are exceedingly painful to the touch, and the stout plates of some of the Goniasters must form admirable organs of protection. The power of restoring lost or injured parts is one of the most remarkable points in the Echinoderm organization (see Case 6).
Echinoderms are of great geological age, and were very abundant in earlier periods of the world’s history. Two groups (the Blastoids and Cystids) have completely disappeared, and the Stalked Crinoids (Lily-Encrinites) are far less common than they used to be. Echinoderms are now found in all seas, and extend to great depths of ocean; many of the species have exceedingly wide areas of distribution, and most are characterized by their gregarious habits, a large number of specimens of a single species being generally obtained by the dredge. They are most abundant in the tropical seas.
Most Echinoderms lay their eggs in the water, where the larvæ 111are developed and swim about freely; but in a few (Hemiaster, Ophiacantha vivipara, and others) the young do not pass through any metamorphosis, for the eggs are placed in special pouches of the body of the parent, in which they are hatched. The free-swimming larvæ of the other Echinoderms pass through a series of remarkable changes (Figs. 4 and 5); these are illustrated by the twelve models of various forms of larvæ exhibited in Case 36; in Case 35 is a set of models showing in detail the changes undergone by a single species (Asterina gibbosa). A portion only of the body of the larva is converted into the substance of the perfect animal; the rest is either absorbed by the growing animal, or shrivels up and disappears.
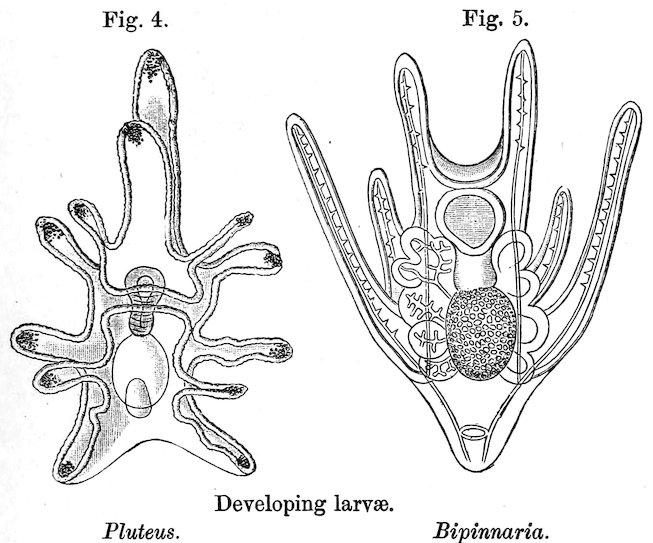
Below the twelve models in Case 36 may be seen a representation of three stages in the history of the Feather-star (Antedon bifida). The larvæ of this Echinoderm are not free, but are attached by a stalk (Fig. 6); in the common Feather-star and other Comatulidæ the stalk is found during larval stages only; in others, such as Pentacrinus, it persists throughout life.
The presence or absence of this stalk has been taken as the first character of importance in the classification of Echinoderma which may be divided into two groups:—
A. Pelmatozoa,[24] or Echinoderms provided with a stalk throughout 112life or in the larval stages only. To this group belong the Crinoidea, and the extinct Blastoidea, and Cystidea.
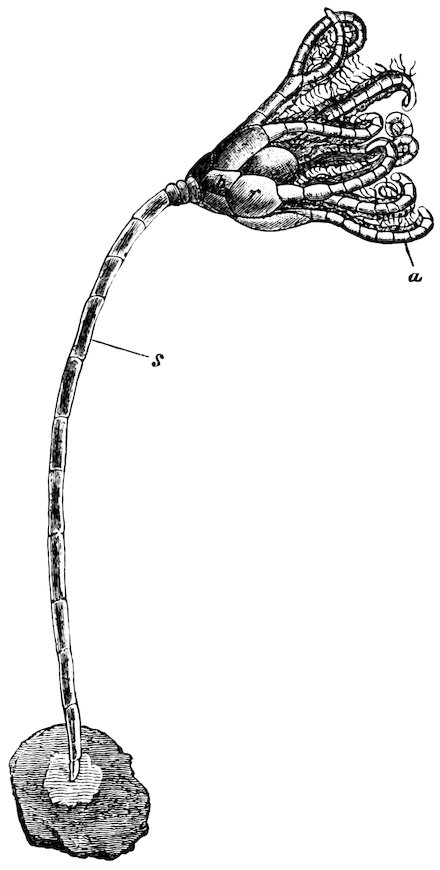
Fig. 6.
Pentacrinoid stage of Antedon rosacea.
a, arms; b, basals; r, radials; s, stalk.
B. Echinozoa, or Echinoderms without stalks at any time of their existence. To this group belong the Asteroidea, Ophiuroidea, Echinoidea, and Holothurioidea.
Crinoidea.—This Order may be described as stalked, globular, or cup-shaped Echinoderms, in which the oral surface of the calyx or disk looks upwards, and in which five jointed and generally branched rays arise from the central disk. Their joints have jointed 113pinnules at their sides, and the sucking-feet have the form of tentacles.
The stalked representatives of this Order are placed on tables and brackets near the south door, and are worthy of being particularly noticed for their fine preservation, size, and beauty. The largest specimen of Pentacrinus decorus was taken on a telegraph-wire, to the covering of which the stalk of the Crinoid is still attached. Metacrinus is a more lately discovered genus, which appears to be confined to the eastern seas.
A few dried unstalked Crinoids are shown in Table-case 1; these show the leading modifications of structure in the two great genera Antedon and Actinometra.
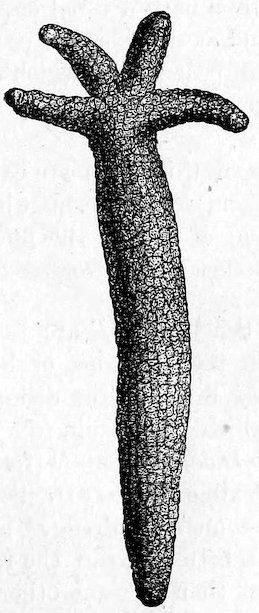
Fig. 7.
Comet form of Linckia.
Asteroidea.—This Order comprises Echinoderms with a depressed body of pentagonal or star-like shape, to the ventral surface of which the ambulacral feet are confined. The rays are more or less elongate movable arms, with skeletal structures, which consist of transversely arranged, paired, calcareous plates, articulated with 114each other like vertebræ, the series extending from the mouth to the end of the arms. The groove in which the ambulacral feet are arranged is uncovered.
Typical specimens of this Order are exhibited in Cases 2 & 3, in which the great variety of form in the genus Asterias and beautiful examples of Acanthaster are shown. Cases 6 & 7 contain specimens illustrating the curious habit of self-mutilation possessed by so many Echinoderms; among Starfishes, and notably in the genus Linckia, the single arms separated from the disk are able to develop a fresh disk and arms, and so to multiply the species. Cases 9–11 contain fine series of Oreaster.
Ophiuroidea, or “Brittle-stars.”—These Echinoderms appear to resemble the ordinary Starfish[25]; but they differ in having the organs of digestion, respiration, and reproduction confined to the disk, the arms having merely the function of locomotor organs. The arms therefore are more slender and cylindrical in form, and are sharply distinct from the disk; the separate joints consist of two central ossicles, which leave only a narrow canal between them, and these are covered above, below, and at the sides by specially developed investing plates; the lateral plates bear spines, which are always comparatively short and delicate, as compared with the spines found at the sides of the arm in starfishes.
The principal types of this Order are exhibited in Cases 17–22; the most exquisite of them are the forms whose arms are divided and subdivided till they end at last in the finest threads, as in Astrophyton, the so-called Basket-fish or Gorgon’s heads.
Echinoidea, or “Sea-Urchins,” are Echinoderms in which the rays are not free, as in the Starfishes or Brittle-stars, but unite to form a compact, spherical, heart- or disk-shaped test; this test is covered with spines, which may attain to a great length, as is shown in the fine example of Diadema saxatile from the Andaman Islands; some of the tests are flexible and very fragile. Owing to the quantity of specimens that are sometimes dredged at one spot, the naturalist has been able to gain a better idea of the range of variation in the species of Echinoderms than in some other divisions of the Animal Kingdom; an instructive series, showing the variations of Echinometra lucunter, is shown in Case 28.
115The genus Hemiaster offers an example of an Echinoderm in which the eggs are laid in special pouches; the hinder ambulacra are deepened to form pits, which are guarded by specially elongated spines (see Case 34); in these pits the young pass through all the stages of their development.
The minute structure of the spines of Sea-Urchins is illustrated by a series of figures on the wall.
The Holothurioidea, or Sea-Cucumbers, form the last order of Echinoderms. Their body, as indicated by their English name, is elongate, subcylindrical, with a more or less flexible integument, according to the extent of the reduction of the calcareous skeleton; the mouth is at one end of the body and surrounded by tentacles, the vent at the opposite end.
As these animals cannot be shown in a dried state, some of them, preserved in spirit, are placed in Wall-Case IV. According as they have or have not the sucking-feet of the Echinoderma, they are ordinarily divided into the Pedata and the Apoda; the latter are represented by Synapta, which may attain to a great length, and by Chiridota; the Pedata are illustrated by the genera Cucumaria, Psolus, and Holothuria. Deep-sea investigations have revealed the existence of another group of specially modified Holothurians—the Elasipoda; these are remarkable for their well-marked bilateral symmetry and the distinctness between the dorsal and ventral portions of the body; the prominent processes on the dorsal surface are not contractile.
An exhibition of some interest is to be found in a Table-Case against the wall, in which there are various specimens of the edible Holothurians—trepang or bêche-de-mer; these were all bought in the market at Canton, and may be taken to be typical of the kinds offered for sale in various eastern countries.
By the name “Worms,” people commonly indicate a number of different forms whose relations with one another are by no means so close as those of a Holothurian and a Crinoid, or a Mussel and an Octopus. There are not, indeed, any common characters by the possession of which the worm-like animals can at once be distinguished from other animals. We take the divisions, examples of which are here represented, either by drawings, models, or specimens preserved in spirit separately.
The groups referred to may be enumerated as follows:—
| Platyhelmia. | Turbellaria. |
| Trematoda. | |
| Cestoda. | |
| Nemertinea. | |
| Nematoidea. | |
| Chætopoda. |
Platyhelmia, or Flat-Worms.—These form the lowest and simplest division of the group.
The parasitic Platyhelmia—the Tapeworms (Cestoda) and the Flukes (Trematoda)—occupy Case I.; the life history of the common Tapeworm (Tænia solium) is shown by the aid of models and figures. A model of the anterior end of the common Tapeworm shows the four suckers and the crown of hooks; the unjointed neck is followed by the joints (proglottids), which increase in size the farther they are from the neck. Several entire specimens of Tænia follow, showing the size of the whole worm and the form of its joints. The structure of the body is shown in the models of two joints. The growth and development of the Tapeworm is dependent on a migration or a change of the hosts which it inhabits in the various stages of its life; and although the different kinds of Tapeworm differ from each other somewhat in certain details of their migration and development, their life history exhibits, on the whole, the same 117events which we find in Tænia solium, a common Tapeworm of man in Northern Europe. This worm is matured in the intestines of man; its final joints consist merely of fertilized ova which have already passed through the earlier stages of development; when the joints are detached and discharged, their contents escape in the form of embryos contained in a thick chitinous shell. If these are now swallowed by a pig, the shell is digested by the gastric juices of the new host, and a rounded embryo, which is provided with three pairs of hooks, is set free; by means of these hooks the guest makes its way through the wall of the stomach or intestine, and finally settles down in the muscles of its host. The embryo now loses its hooks, and gradually acquires a bladder-like form, the central cavity of which is filled with fluid. This bladder-worm (Cysticercus) has its outer wall pushed inwards at the anterior end, and on this hooks and suckers become developed. We have now a narrow head and neck with an attached bladder, the head being at this time hollow. If during the long time that these bladder-worms remain alive, the pig is killed for food, its flesh is found to be “measly”; if it is afterwards insufficiently cooked and eaten, the worms are conveyed into the human stomach. Here the bladder-like termination becomes absorbed, and, the neck beginning to grow, we have the commencement of the form from which we started, and the completion of that “vicious circle” which is so curious a characteristic of many forms of parasitic life.
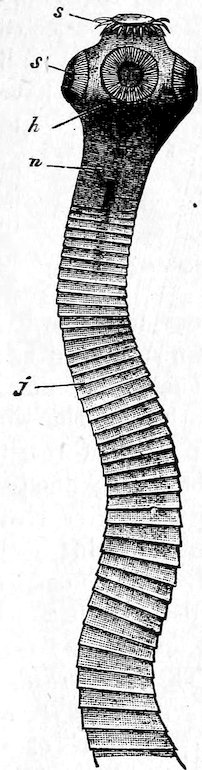
Fig. 8.
Tænia solium: showing the head (h) with its suckers (s′) and crown of hooks (s), the unjointed neck (n), and a few of the succeeding joints (j).
In other Tapeworms the cyst may be more complicated than that in the pig, as, for example, the form found in the sheep’s brain or the liver of the horse.
Of the other Cestode parasites mention should specially be made of those of Fishes; the vulgar notion that the parasites of these 118animals are dangerous to man has been shown to be entirely erroneous.

Fig. 9.
Limnæa truncatula.
The Flukes infest animals of all kinds; that which is most dangerous to sheep, and the cause of much pecuniary loss (Distoma hepaticum), is selected here as a type; its structure is shown by a large model, and its life history by a series of diagrams (Figs. 10–13). Here, again, we have a creature which infests two hosts. If the larvæ which escape from the sheep fall on wet ground in or near a pool, they make their way to a small pond-snail (Limnæa truncatula, Fig. 9), into the lung-chamber of which they bore their way. On leaving them the larva may be, and is, too frequently, eaten by a sheep, and makes its way into the liver of that animal, where it causes the disease known as the “liver rot.”
The damage done by the liver-fluke may be imagined from the fact that in the winter of 1879–80 no less than three millions of sheep died of rot in the United Kingdom; this heavy loss is no doubt largely due to the immense number of eggs to which a single fluke may give rise. It has been estimated that every fluke may produce, during its life, several thousands of eggs; and in one case Prof. A. P. Thomas found as many as 7,400,000 eggs in the gall-bladder of a sheep which was suffering from rot, and which, at that time, had in its liver about 200 flukes.
The non-parasitic Flat-worms are shown, magnified, in the upper parts of Cases I. & II. The Turbellaria proper, without any or with a simple or a branched intestine, but without a vent, are represented by Convoluta and Thysanozoon: the general structure is shown by a diagram in Case II., which is here reproduced (Fig. 14). Planaria, Thysanozoon, and Bipalium serve to illustrate the forms of members of this group.
The Nemertine Worms (Nemertinea), with a straight intestine, with a vent, and with a proboscis, may attain to a very considerable length; Carinella and Lineus are represented by large figures, and various species are shown in spirit. These forms, which used to be very unsatisfactory to exhibit, on account of the great difficulty of preserving them complete and uninjured, are now, with improved methods, very satisfactorily shown, as the specimens purchased from the Marine Biological Laboratory at Plymouth prove.
Stages in the life history of the Fluke.
Fig. 10. Egg of Fluke, showing the operculum and the contained yolk-spheres. Magnified 340 diams.
Fig. 11. An embryo forcing its way by its boring-papilla (p) into the wall of the lung of a Snail (e.p.) Magnified about 340 diams.
Fig. 12. A young Rédia (natural size, ½ millimetre or ¹⁄₅₀ inch): pl., pharynx; g, contained germs; p, characteristic posterior processes of the Rédia.
Fig. 13. Free-swimming Cercaria, before the commencement of the formation of the cyst. Magnified 100 diams.
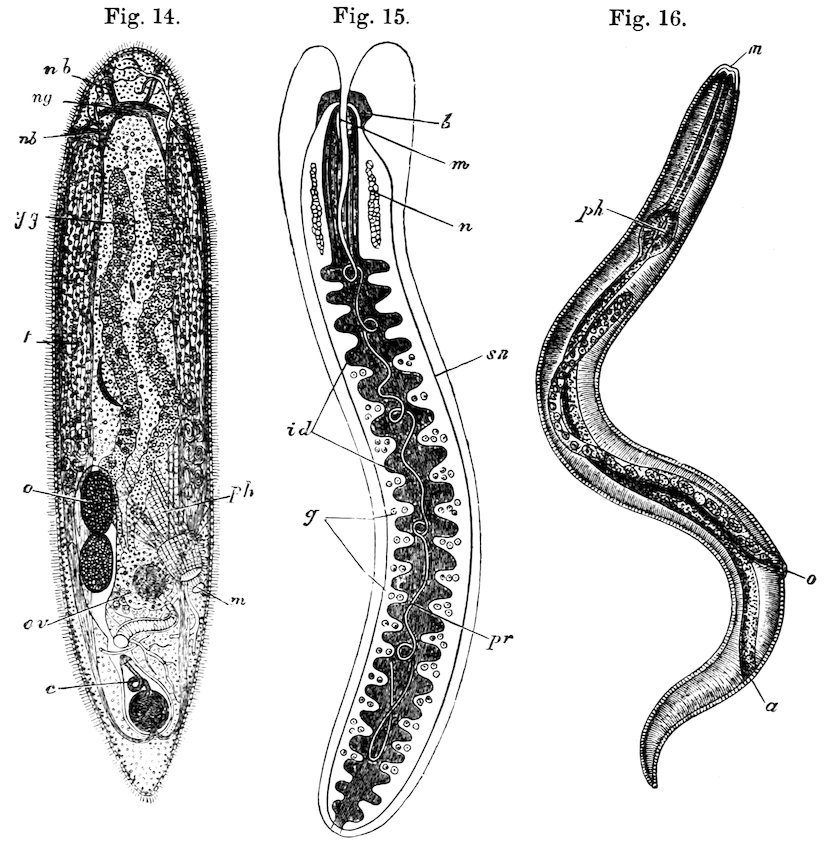
Fig. 14. Diagram of the structure of a Turbellarian: ng, nerve- (cerebral) ganglia; nb, nerve-branches; yg, yolk-glands; t, testis; o, ova; ov, ovary; c, cirrus; m, mouth; ph, pharynx.
Fig. 15. Diagram of a Nemertine: b, brain; m, mouth; n, renal organs; id, diverticula of intestine; g, gonads; sn, side nerve-trunk; pr, proboscis in its dorsal sheath.
Fig. 16. Diagram of the structure of a Nematoid; m, mouth; ph, pharynx; a, anus; o, orifice of genital tube.
Nematodes (Thread-Worms or Round-Worms).—These are for the most part parasitic, and infest plants as well as animals; the common Round-Worms living parasitically in man (Ascaris, Stronaylus, Trichocephalus) belong to this Order. Sometimes they are parasitic in their early stages and later live a free life—such are Gordius and Mermis. A specimen of a Mantid is exhibited from which half the body of the infesting Gordius has already protruded (Fig. 17). One of the most remarkable Gordii is the great elongated 121G. fulgur, or “Lightning Snake,” from Celebes. Another very large Nematode is the so-called Guinea-worm, or Dracunculus medinensis, which is found beneath the skin of the leg; it is very possible that this worm was the cause of the illness which afflicted the Israelites in their journey through the desert from Egypt to the Promised Land.
Fig. 17.
Gordius escaping from a Mantid.

Fig. 18.
Figure of Trichina spiralis, showing the worms encysted in muscle.
Of all Nematodes the most dangerous to man is the small worm which is known as Trichina spiralis (Fig. 18); a series of models are 122shown which give a good idea of the structure of the female and the smaller male. The young make their way through the walls of the stomach of their host, and encyst themselves among its muscles: a piece of a sternothyroid muscle is shown, taken from a man in whose body it was calculated there were forty millions of encysted Trichinæ.
Other Nematodes infesting man, such as Filaria sanguinis hominis, are too small for exhibition.
Plants are not free from the attacks of Nematodes, and examples are shown, accompanied by an illustrating figure, of the Ear-cockle gall of wheat; this gall is due to the injuries inflicted by a minute Thread-worm—Tylenchus tritici. Wheat is, of course, by no means the only cultivated plant that is attacked by these minute worms; the history of most has, however, still to be made out.
Holding a somewhat uncertain position in relation to the Round-worms are the parasitic Acanthocephali (Thorn-headed Worms) and the free-swimming Chætognatha, or Bristle-jawed Worms; examples of both of these groups are shown, together with diagrams illustrative of their general structure.
Annulata or Chætopoda.—So-called because consisting of a series of rings, and being provided with chætæ or bristles; they are to be associated with the Arthropoda, under the one head “Appendiculata,” a better name than “Articulata,” since Cuvier did not include worms in his group. The creatures that are most familiarly called worms are to be found in Case III.; here are a few examples of the numerous kinds of worms that are found living freely in the sea, of earth and freshwater Worms, and of Leeches. All these worms are distinctly characterized by the fact that they consist of a number of definite rings (somites), whence they have been called Annulata. The marine Worm and the Earthworm differ from the Leech in that these rings are provided with chætæ or bristles, of which there are a number in each bundle in the marine, and a few only in the terrestrial or freshwater form: hence the marine Worms are called Polychæta and the latter Oligochæta.
The former are divisible into two great groups. There are those that are free-swimming and are able to forage for themselves, such as the lovely Sea-mouse (Aphrodite aculeata), the large Eunice gigantea, the common Nereis pelagica, or the exquisitely coloured Chloeia flava. Others live a more retired life, dwelling in tubes, which they fashion for themselves; they lead either a solitary or a social life. Here we have examples of Sabella, Sabellaria, Serpula; 123a number of forms of worm-tubes, showing their great variety and beauty (see especially the delicate Filograna), are to be seen in the small Table-cases placed against the north wall of the Gallery. Attention should be especially directed to Mr. A. T. Watson’s beautiful preparations of Terebella littoralis. We give a figure (Fig. 19) after a drawing by that gentleman of the home of Panthalis oerstedi, the tube-forming habits of which have been carefully observed by him.
Fig. 19.
Home of Panthalis oerstedi.
Fig. 20.
Section across the body of an earth-worm to show the disposition of the more important organs; the body wall (w) consists of dermis, circular, and longitudinal muscles; the body cavity is divided by membranes (c) into a series of chambers, in each of which opens the mouth of a coiled nephridium (n). The axis of the cavity is occupied by the intestine (i); above and below it is a longer blood-vessel (v), and below it is also the central nerve-cord (nc).
124The Oligochæta are represented by the common Earthworm, the influence of which in the formation of mould and in the general ploughing of the soil was carefully investigated by Mr. Darwin; and by the little Tubifex rivulorum (Bloodworm), which owes both its red colour and its ability to dwell in mud, which is so poor in oxygen as to be unfit for respiration, to the same chemical compound as that which gives the red colour to our blood and carries the oxygen of respiration all over the body.
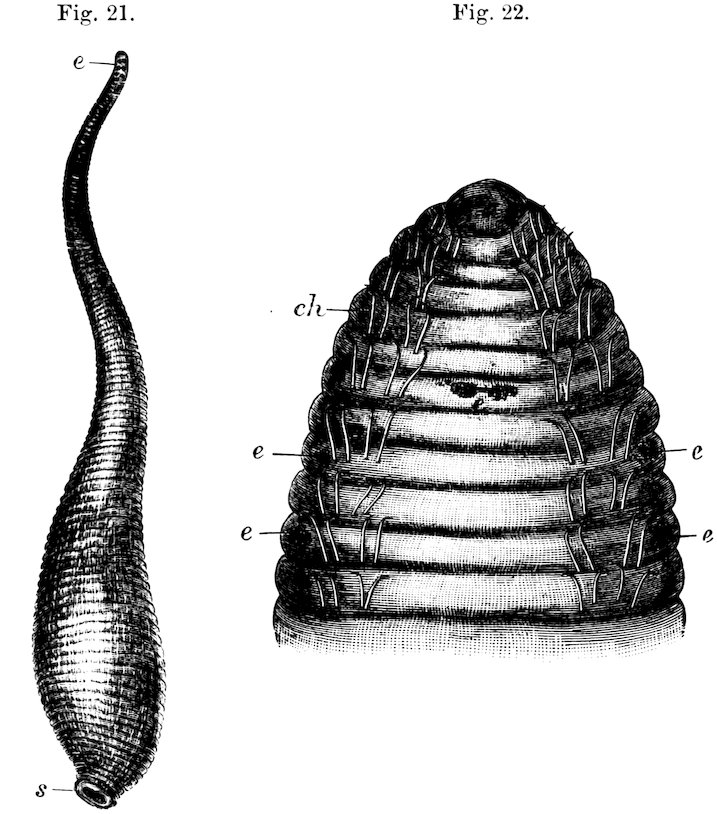
Acanthobdella: e, eyes; ch, chætæ; s, sucker.
The Hirudinea, or Leeches, are often said to be distinguished from the Chætopoda by the absence of bristles, but, as a fact, Acanthobdella (Figs. 21 and 22) has very well marked bristles. They always have a sucker at the hinder end of the body by which they are attached to their prey; they are found in fresh water (Piscicola), on sea-fishes (as Pontobdella), or in moist places, as the Leech (Hirudo). The last-named has three jaws, armed with as many 125as ninety denticles. Trochetia subviridis (Land-Leech) is a species which is found rarely and sporadically in England.
The Myzostomaria form a division of Polychæta all the members of which live parasitically on Crinoids, and otherwise present great differences in their habits. Some move about freely on the Crinoids they infest, others are more sluggish and rarely move, others produce galls or cysts on their host, and yet others are internal parasites, and live in the alimentary canal. It is of interest to note that there are corresponding degrees of difference between the young and old specimens of the different groups of species.
The general organisation of Myzostomaria is shown in the accompanying figure (Fig. 23) in which the dorsal wall of the body is supposed to be transparent so as to allow of the chief internal organs being seen.
Fig. 23.
Diagram of Myzostomum to show the general form of the body and the marginal extensile cirri (c); within these and on the ventral surface are four pairs of suckers, and more internally five pairs of appendages each bearing two hooks; the proboscis (p), the digestive tract and its ramifications, and the reproductive organs are outlined as if seen through a transparent wall; a, anus.
126The last group of Worms here represented is that of the Gephyrea; with the advance of our knowledge it is probable that they will be found to be more intimately allied to the Annulata than is now generally supposed; it will be seen indeed that Echiurus has bristles at its hinder end; Sipunculus is the best known representative of the unarmed Gephyrea; Bonellia is interesting both from the fact that it owes its green colour to a matter closely resembling the chlorophyll of green plants, and from the possession by the female of a proboscis, which is protruded from the hole in the rock occupied by the worm: the male is very much smaller than the female, and is not nearly so well developed. Owing to the mode of lighting the Gallery, the visitor may have to shift his position several times before gaining a good view of the whole length of the proboscis.
1. A framed series of photographs, illustrating different kinds of radulæ, is placed on the east wall of the gallery.
2. From Woodward’s ‘Manual of the Mollusca,’ published by Lockwood & Son.
3. From ‘The Cambridge Natural History,’ Messrs. Macmillan & Co.
4. From the Greek: pteron, wing, and pous, foot.
5. From the Greek: scaphe, a small boat, and pous, a foot—the foot of some Scaphopods being somewhat pointed like the prow of a vessel.
6. The term Lamellibranchia is used instead of Pelecypoda in deference to the wish of Professor Lankester.—E. A. S.
7. From the ‘Encyclopædia Britannica.’ Messrs. A. & C. Black.
8. From ‘The Cambridge Natural History.’ Messrs. Macmillan & Co.
9. From the Greek: kephale, head, and pous, foot.
10. Zoon, animal; oikos, house.
11. Lophos, plume; pherein, to bear.
12. Polus, many; zoon, animal.
13. Bryon, moss.
14. Ektos, outside; proktos, vent.
15. Cheilos, lip; stoma, mouth.
16. Gumnos, naked; laimos, throat.
17. Ktenos, of a comb.
18. Phulassein, to guard; laimos, throat.
19. Kuklos, circle.
20. Entos, inside; proktos, vent.
21. Vibraculum, a bristle.
22. Pyrosoma—pur, fire; soma, body.
23. It should be mentioned that one high authority, Prof. W. K. Brooks, does not regard the life history of Salpa as an example of alternation of generations, but considers the solitary Salpa to be, not asexual, but a female which produces a chain of males; but it is impossible to enter into a difficult question of controversy here.
24. From the Greek pelma = a stalk.
25. The Asteroidea or Ophiuroidea may be counted under the name Stelliformia.